Department of Medical Genetics, and Cambridge Institute for Medical Research (CIMR), University of Cambridge, Cambridge, UK.
UK Dementia Research Institute, Cambridge Institute for Medical Research (CIMR), University of Cambridge, Cambridge, UK.
Nat Cell Biol. 2024 Oct;26(10):1691-1699. doi: 10.1038/s41556-024-01488-7. Epub 2024 Aug 29.
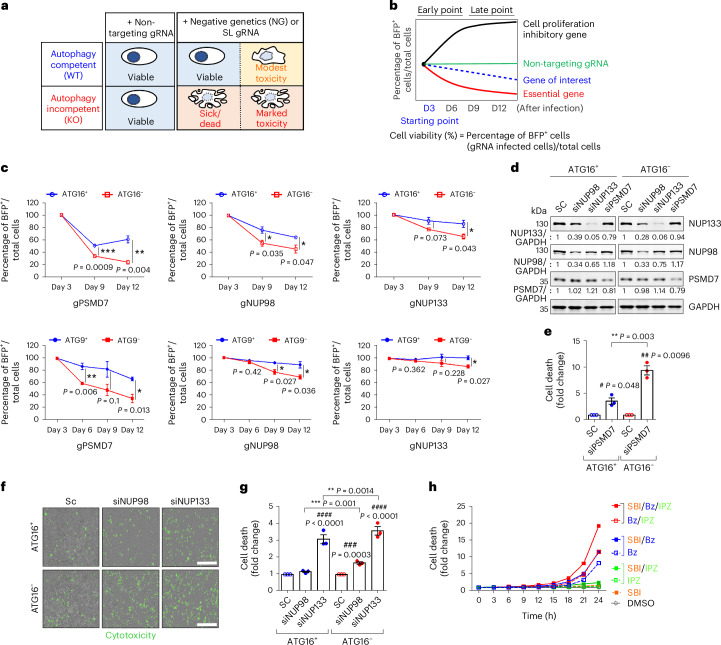
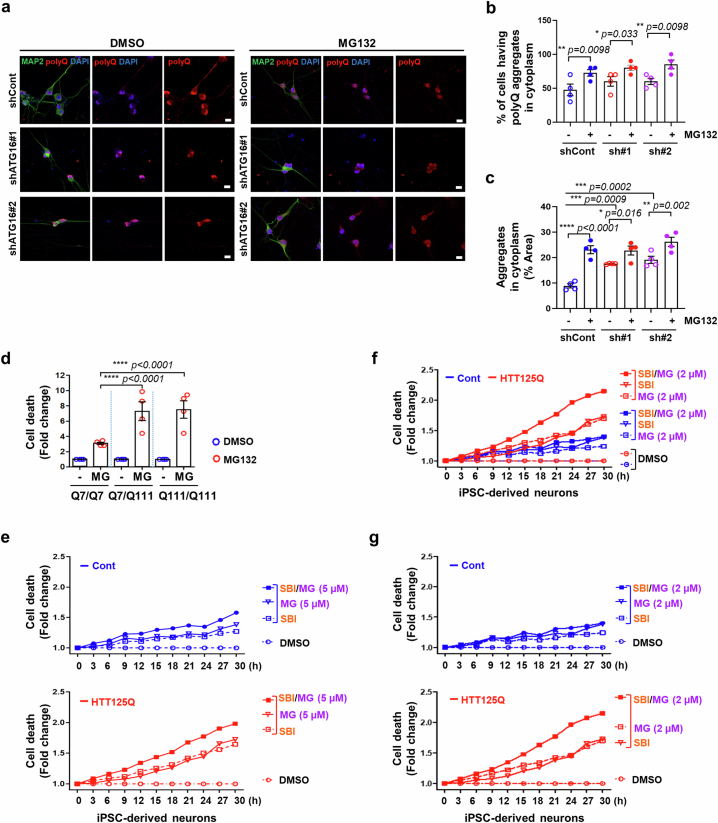
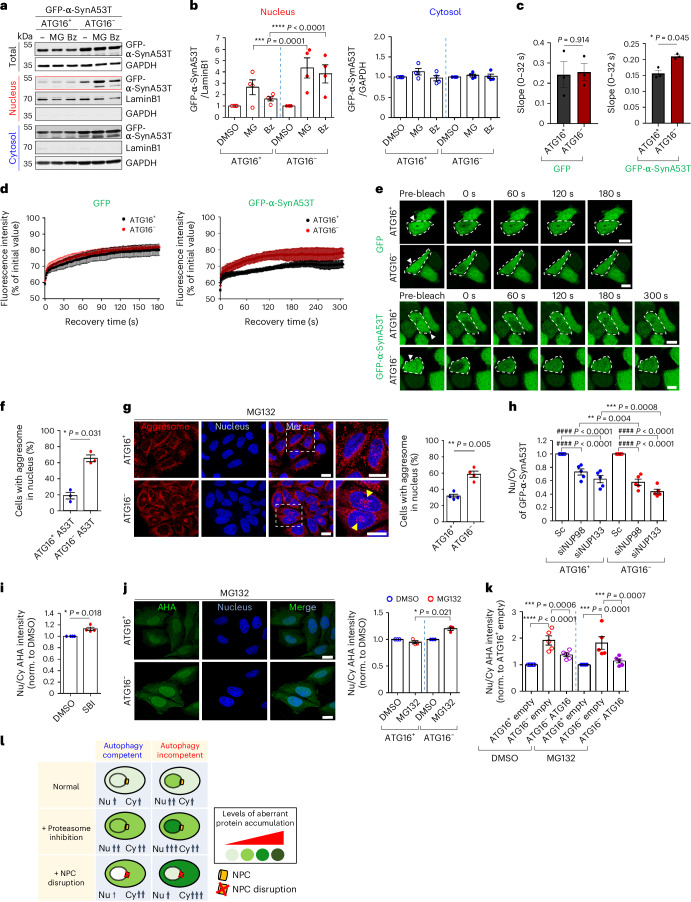
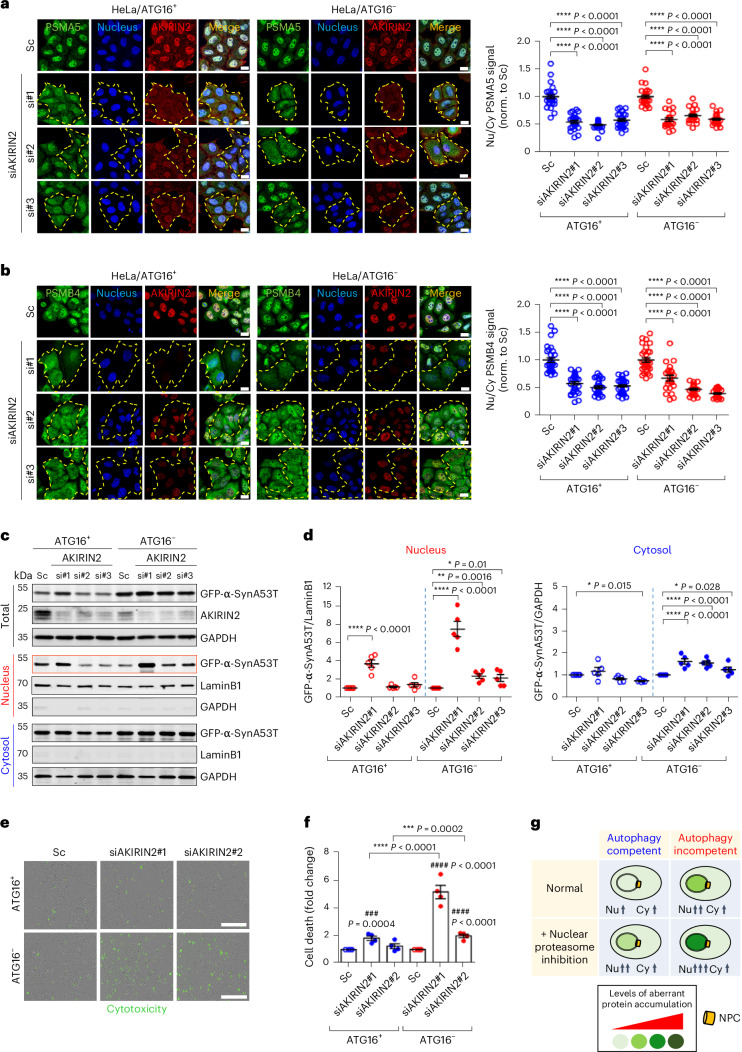
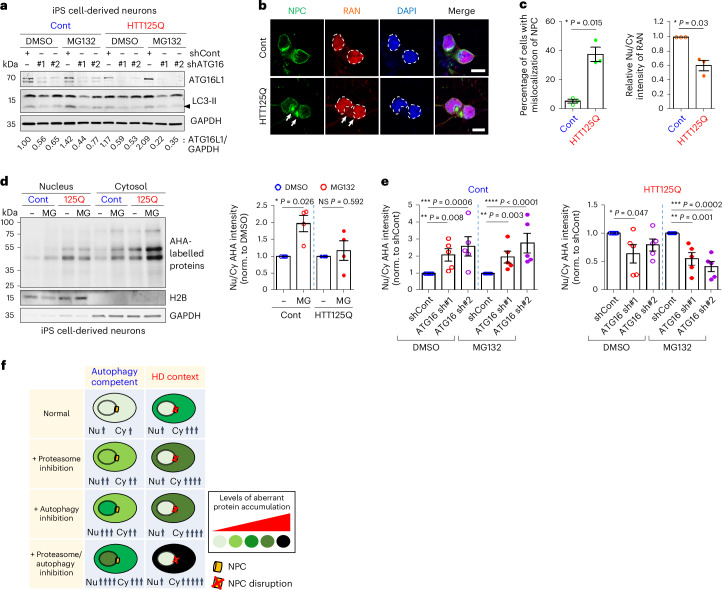
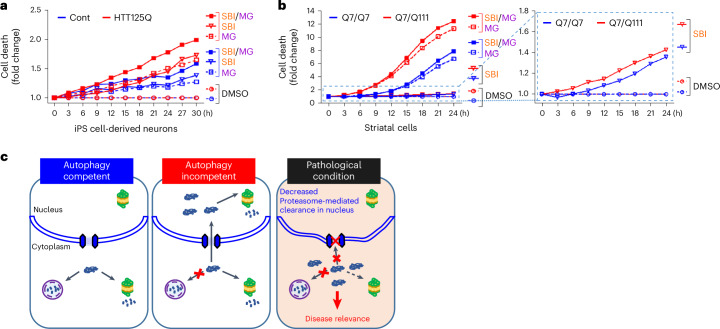
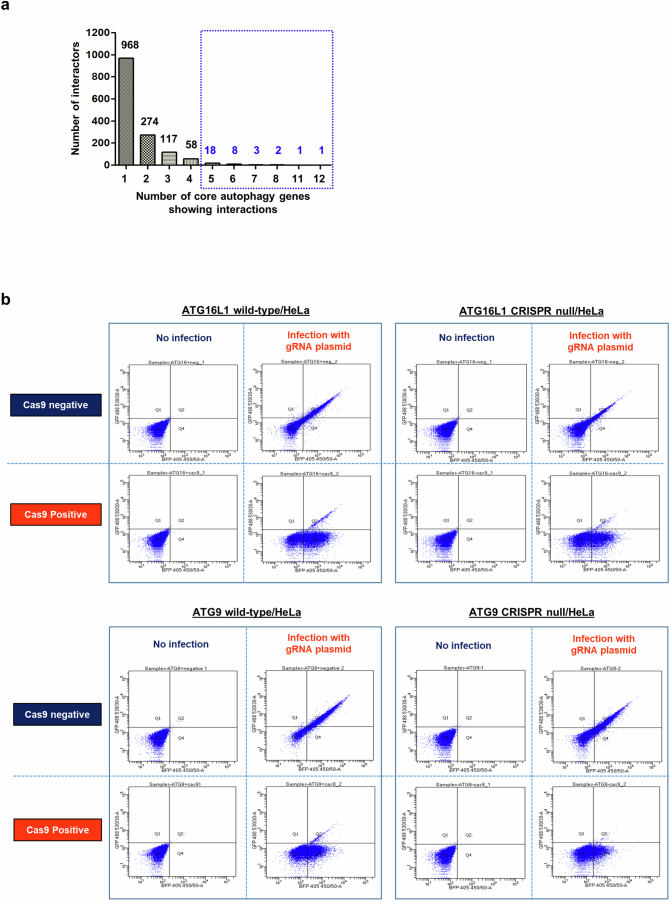
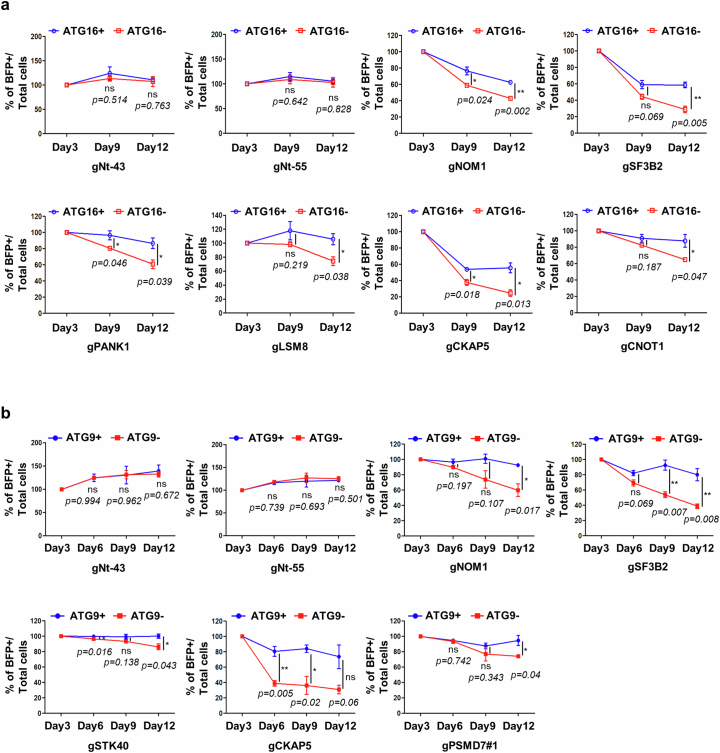
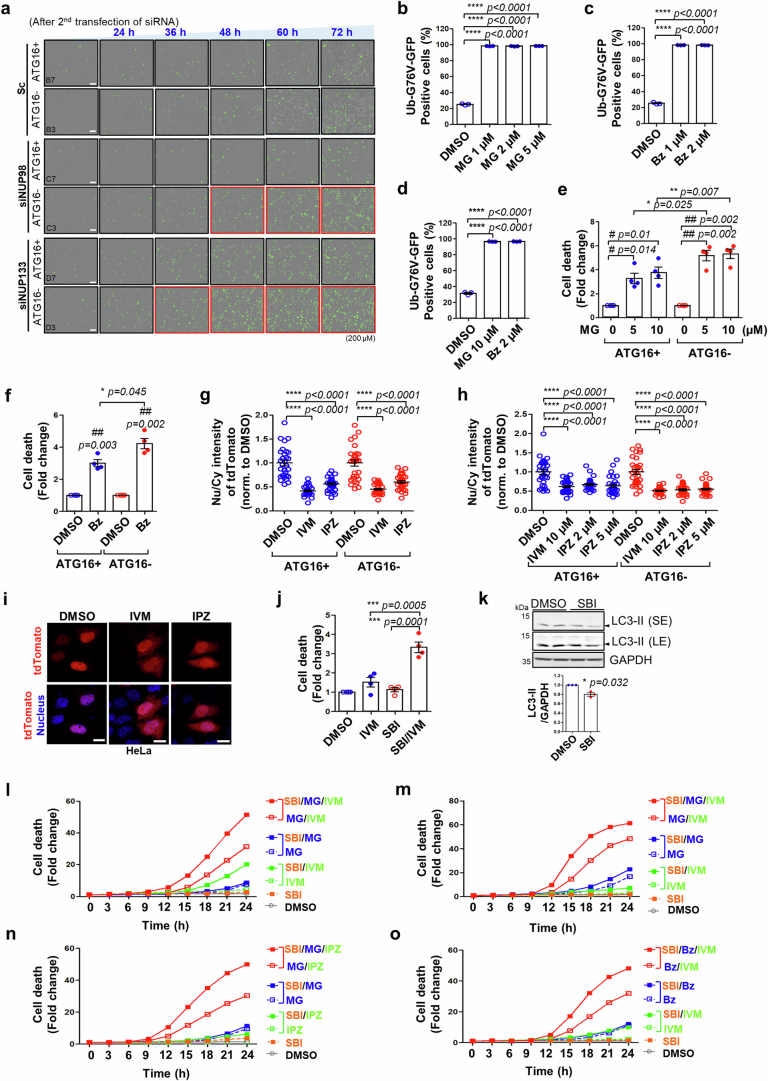
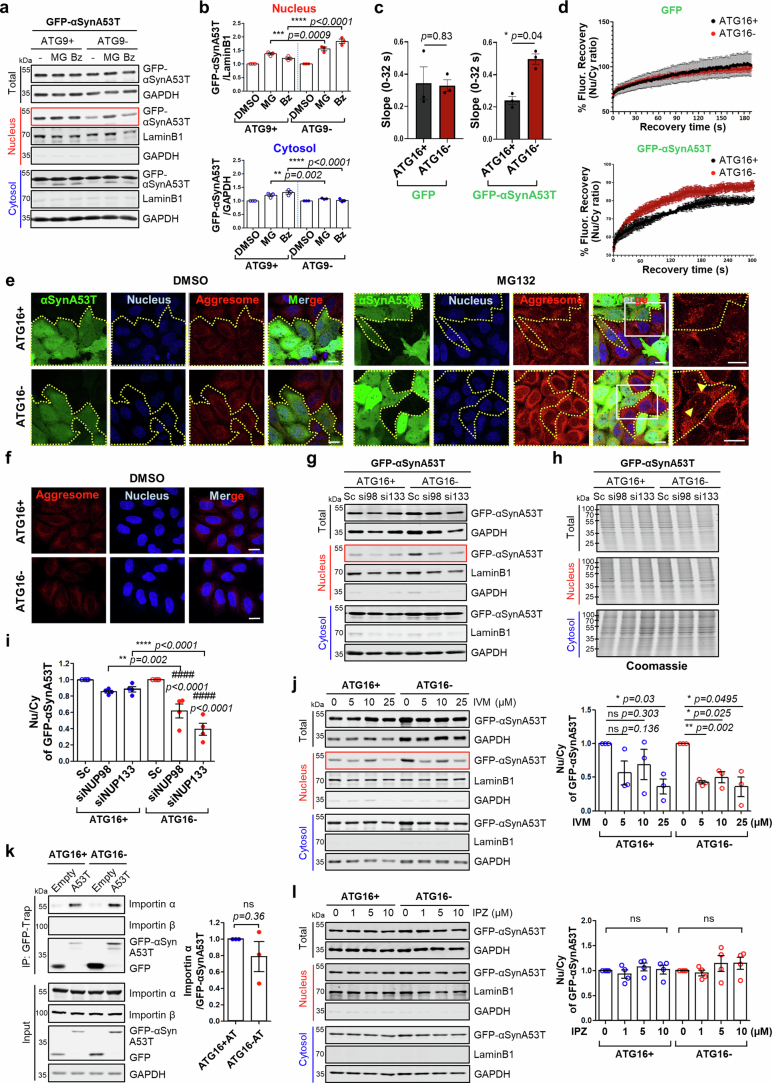
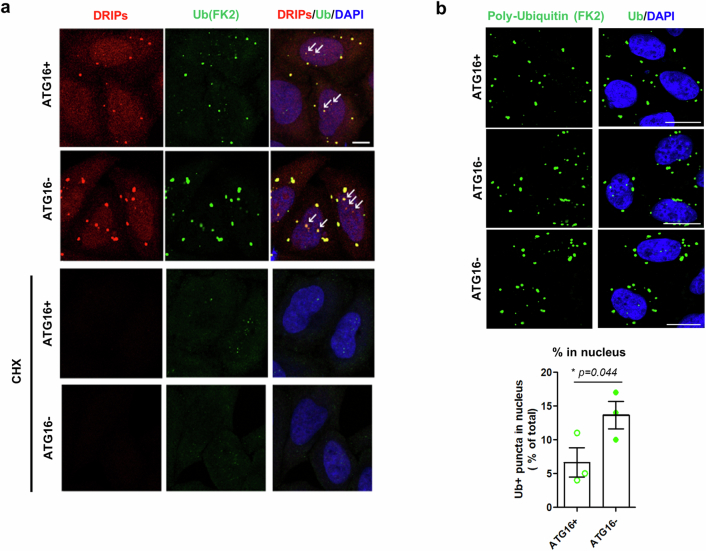
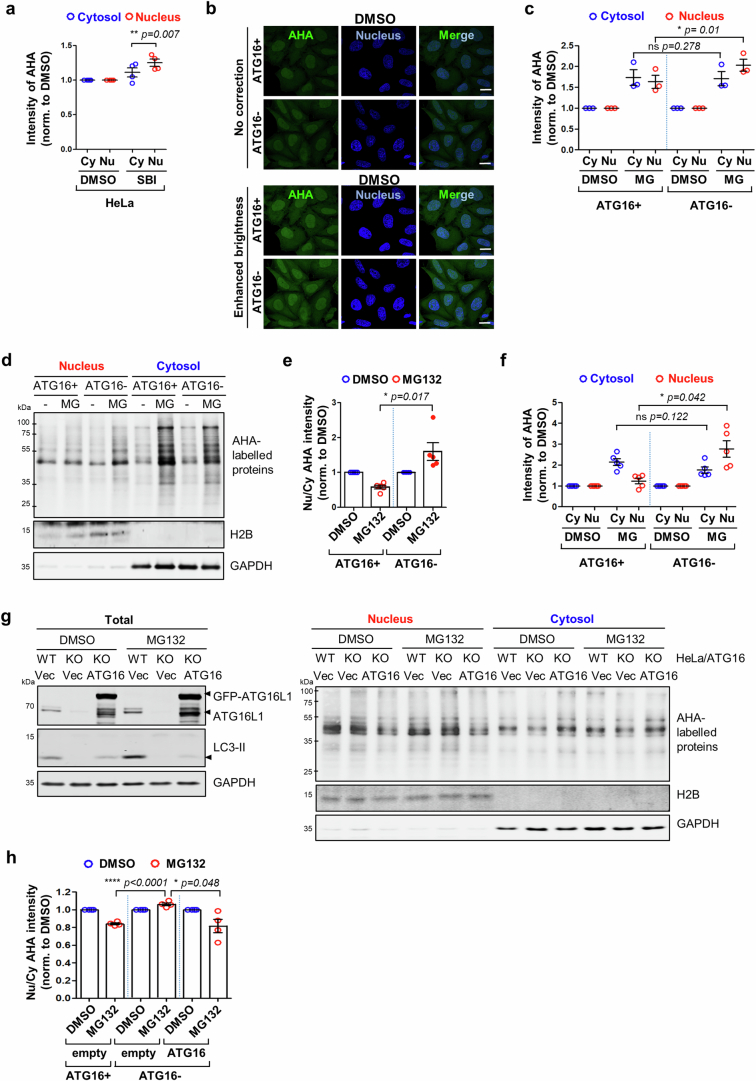
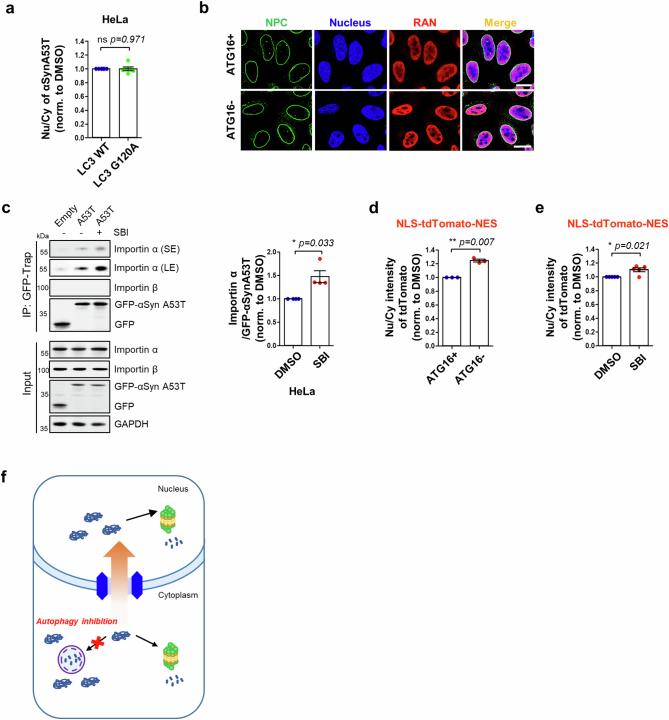
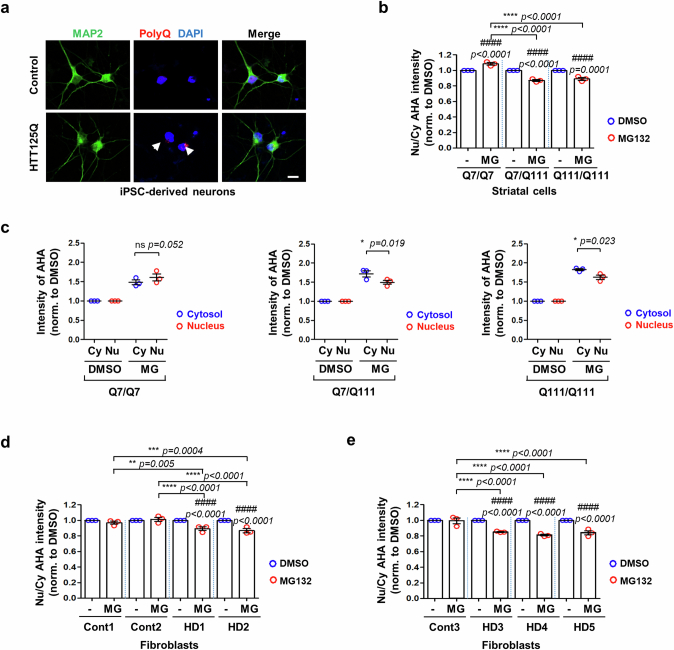
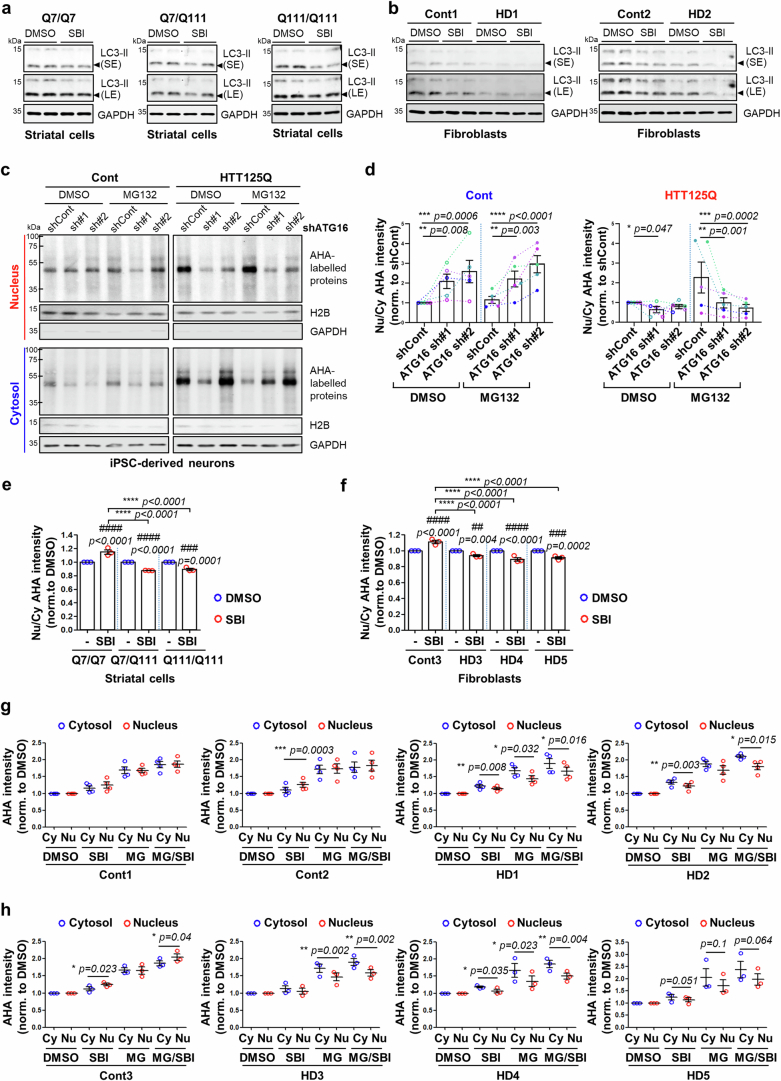
Autophagy is a conserved pathway where cytoplasmic contents are engulfed by autophagosomes, which then fuse with lysosomes enabling their degradation. Mutations in core autophagy genes cause neurological conditions, and autophagy defects are seen in neurodegenerative diseases such as Parkinson's disease and Huntington's disease. Thus, we have sought to understand the cellular pathway perturbations that autophagy-perturbed cells are vulnerable to by seeking negative genetic interactions such as synthetic lethality in autophagy-null human cells using available data from yeast screens. These revealed that loss of proteasome and nuclear pore complex components cause synergistic viability changes akin to synthetic fitness loss in autophagy-null cells. This can be attributed to the cytoplasm-to-nuclear transport of proteins during autophagy deficiency and subsequent degradation of these erstwhile cytoplasmic proteins by nuclear proteasomes. As both autophagy and cytoplasm-to-nuclear transport are defective in Huntington's disease, such cells are more vulnerable to perturbations of proteostasis due to these synthetic interactions.
自噬是一种保守的途径,其中细胞质内容物被自噬体吞噬,然后自噬体与溶酶体融合,使它们被降解。核心自噬基因的突变会导致神经疾病,自噬缺陷也可见于帕金森病和亨廷顿病等神经退行性疾病。因此,我们试图通过寻找负遗传相互作用,如自噬缺失人类细胞中的合成致死性,来了解自噬受到干扰的细胞容易受到哪些细胞通路干扰,利用酵母筛选的现有数据进行研究。这些研究表明,蛋白酶体和核孔复合物成分的缺失会导致协同生存能力的变化,类似于自噬缺失细胞中的合成适应性丧失。这可以归因于自噬缺陷期间细胞质到细胞核的蛋白质转运,以及随后由核蛋白酶体对这些先前细胞质蛋白质的降解。由于亨廷顿病中自噬和细胞质到细胞核的运输都有缺陷,因此这些细胞更容易受到蛋白质稳态失调的影响,这是由于这些合成相互作用所致。