Schipper Louis A, Reeve Jennifer L, Arcus Vickery L, Isson Terry, Prentice Erica J, Williamson Adele, Barnes Andrew D
Te Aka Mātuatua School of Science The University of Waikato Hamilton Aotearoa New Zealand.
Te Aka Mātuatua School of Science The University of Waikato Tauranga Aotearoa New Zealand.
Ecol Evol. 2024 Dec 12;14(12):e70701. doi: 10.1002/ece3.70701. eCollection 2024 Dec.
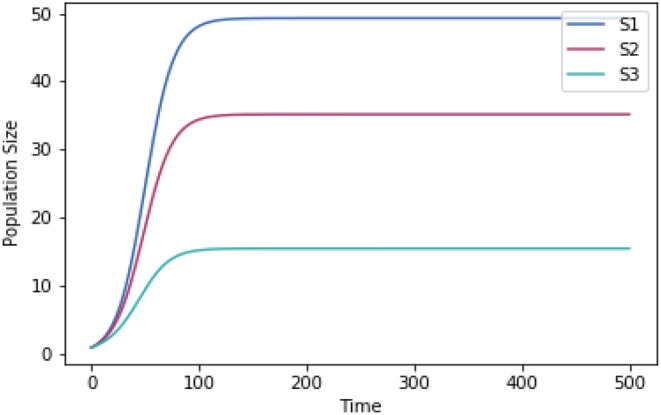
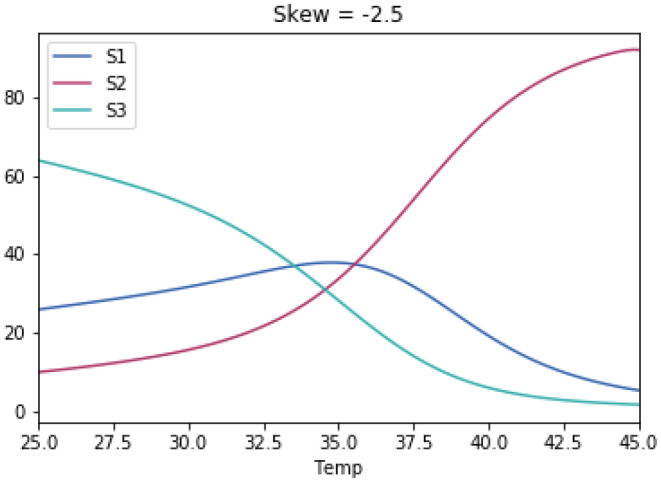
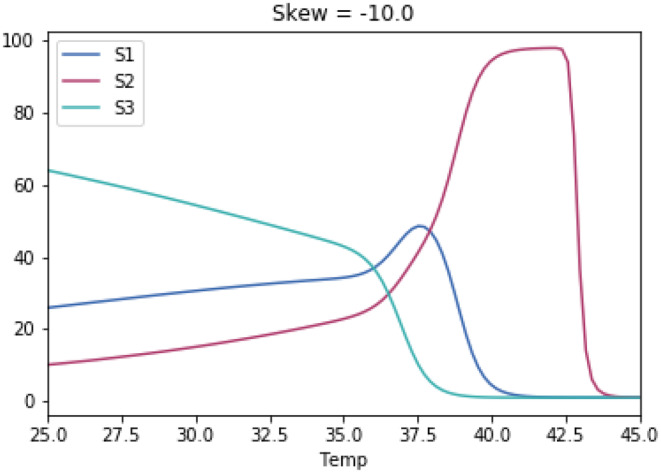
Why does the growth of most life forms exhibit a narrow range of optimal temperatures below 40°C? We hypothesize that the recently identified stable range of oceanic temperatures of ~5 to 37°C for more than two billion years of Earth history tightly constrained the evolution of prokaryotic thermal performance curves to optimal temperatures for growth to less than 40°C. We tested whether competitive mechanisms reproduced the observed upper limits of life's temperature optima using simple Lotka-Volterra models of interspecific competition between organisms with different temperature optima. Model results supported our proposition whereby organisms with temperature optima up to 37°C were most competitive. Model results were highly robust to a wide range of reasonable variations in temperature response curves of modeled species. We further propose that inheritance of prokaryotic genes and subsequent co-evolution with microbial partners may have resulted in eukaryotes also fixing their temperature optima within this narrow temperature range. We hope this hypothesis will motivate considerable discussion and future work to advance our understanding of the remarkable consistency of the temperature dependence of life.
为什么大多数生命形式的生长在低于40°C的狭窄最佳温度范围内表现出来?我们推测,在地球历史超过20亿年的时间里,最近确定的约5至37°C的海洋温度稳定范围,将原核生物热性能曲线的进化紧紧限制在生长的最佳温度低于40°C。我们使用具有不同最佳温度的生物体之间种间竞争的简单洛特卡-沃尔泰拉模型,测试了竞争机制是否重现了观察到的生命最佳温度上限。模型结果支持了我们的观点,即最佳温度高达37°C的生物体最具竞争力。模型结果对建模物种温度响应曲线的广泛合理变化具有高度鲁棒性。我们进一步提出,原核基因的遗传以及随后与微生物伙伴的共同进化,可能导致真核生物也将其最佳温度固定在这个狭窄的温度范围内。我们希望这个假设能引发大量讨论和未来的工作,以推进我们对生命温度依赖性显著一致性的理解。