Genot E, Cleverley S, Henning S, Cantrell D
Lymphocyte Activation Laboratory, Imperial Cancer Research Fund, 44 Lincoln's Inn Fields, London WC2A 3PX, UK.
EMBO J. 1996 Aug 1;15(15):3923-33.
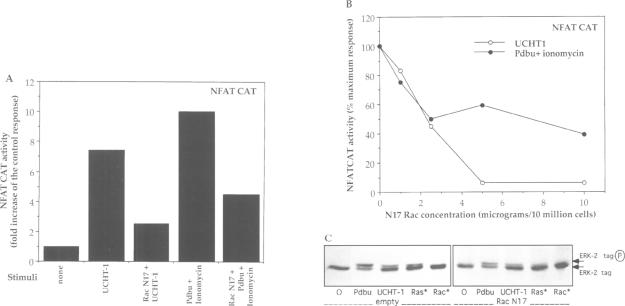
The transcription factor, Nuclear Factor of Activated T cells (NFAT) is a major target for p21ras and calcium signalling pathways in the IL-2 gene and is induced by p21ras signals acting in synergy with calcium/calcineurin signals. One p21ras effector pathway involves the MAP kinase ERK-2, and we have examined its role in NFAT regulation. Expression of dominant negative MAPKK-1 prevents NFAT induction. Constitutively active MAPKK-1 fully activates ERK-2 and the transcription factor Elk-1, but does not substitute for activated p21ras and synergize with calcium/calcineurin signals to induce NFAT. Expression of dominant negative N17Rac also prevents TCR and p21ras activation of NFAT, but without interfering with the ERK-2 pathway. The transcriptional activity of the NFAT binding site is mediated by a complex comprising a member of the NFAT group and AP-1 family proteins. The induction of AP-1 by p21ras also requires Rac-1 function. Activated Rac-1 could mimic activated p21ras to induce AP-1 but not to induce NFAT. Moreover, the combination of activated MAPKK-1 and Rac-1 could not substitute for activated p21ras and synergize with calcium signals to induce NFAT. Thus, p21ras regulation of NFAT in T cells requires the activity of multiple effector pathways including those regulated by MAPKK-1/ERK-2 and Rac-1.
转录因子活化T细胞核因子(NFAT)是白细胞介素-2基因中p21ras和钙信号通路的主要靶点,由p21ras信号与钙/钙调神经磷酸酶信号协同作用诱导产生。一条p21ras效应通路涉及丝裂原活化蛋白激酶ERK-2,我们研究了其在NFAT调控中的作用。显性负性MAPKK-1的表达可阻止NFAT的诱导。组成型活性MAPKK-1可完全激活ERK-2和转录因子Elk-1,但不能替代活化的p21ras,也不能与钙/钙调神经磷酸酶信号协同诱导NFAT。显性负性N17Rac的表达也可阻止TCR和p21ras对NFAT的激活,但不干扰ERK-2通路。NFAT结合位点的转录活性由一个包含NFAT家族成员和AP-1家族蛋白的复合物介导。p21ras对AP-1的诱导也需要Rac-1功能。活化的Rac-1可模拟活化的p21ras诱导AP-1,但不能诱导NFAT。此外,活化的MAPKK-1和Rac-1的组合不能替代活化的p21ras,也不能与钙信号协同诱导NFAT。因此,p21ras对T细胞中NFAT的调控需要多种效应通路的活性,包括由MAPKK-1/ERK-2和Rac-1调控的通路。