Redding K, Seeger M, Payne G S, Fuller R S
Department of Biochemistry, Stanford University, California 94305, USA.
Mol Biol Cell. 1996 Nov;7(11):1667-77. doi: 10.1091/mbc.7.11.1667.
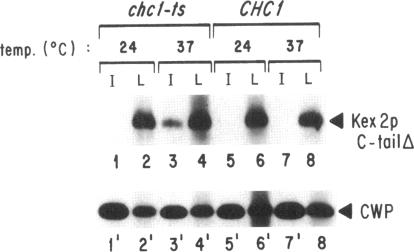
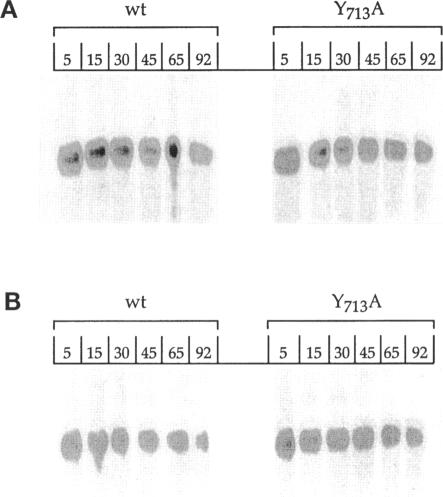
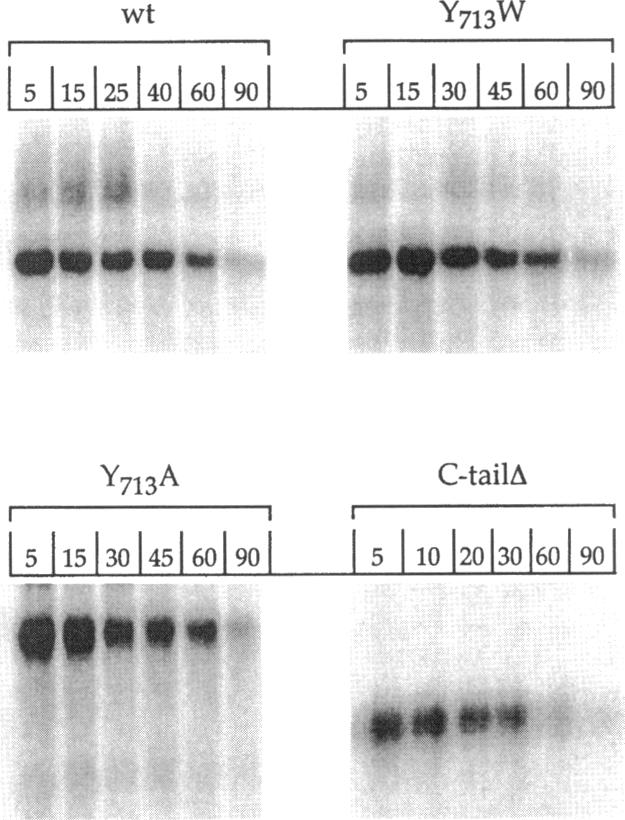
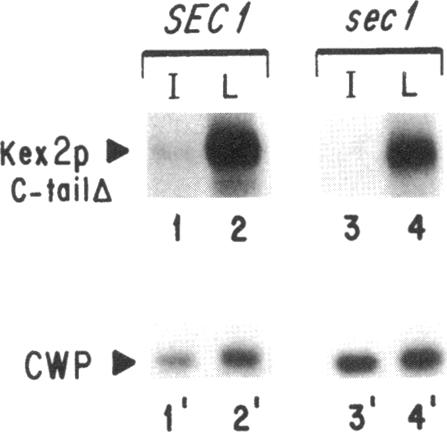
Localization of Kex2 protease (Kex2p) to the yeast trans-Golgi network (TGN) requires a TGN localization signal (TLS) in the Kex2p C-terminal cytosolic tail. Mutation of the TLS accelerates transport of Kex2p to the vacuole by an intracellular (SEC1-independent) pathway. In contrast, inactivation of the clathrin heavy-chain gene CHC1 results in transport of Kex2p and other Golgi membrane proteins to the cell surface. Here, the relationship of the two localization defects was assessed by examining the effects of a temperature-sensitive CHC1 allele on trafficking of wild-type (WT) and TLS mutant forms of Kex2p. Inactivation of clathrin by shifting chc1-ts cells to 37 degrees C caused WT and TLS mutant forms of Kex2p to behave identically. All forms of Kex2p appeared at the plasma membrane within 30-60 min of the temperature shift. TLS mutant forms of Kex2p were stabilized, their half-lives increasing to that of wild-type Kex2p. After inactivation of clathrin heavy chain, vacuolar protease-dependent degradation of all forms of Kex2p was blocked by a sec1 mutation, which is required for secretory vesicle fusion to the plasma membrane, indicating that transport to the cell surface was required for degradation by vacuolar proteolysis. Finally, after clathrin inactivation, all forms of Kex2p were degraded in part by a vacuolar protease-independent pathway. After inactivation of both chc1-ts and sec1-ts, Kex2 was degraded exclusively by this pathway. We conclude that the effects of clathrin inactivation on Kex2p localization are independent of the Kex2p C-terminal cytosolic tail. Although these results neither prove nor rule out a direct interaction between the Kex2 TLS and a clathrin-dependent structure, they do imply that clathrin is required for the intracellular transport of Kex2p TLS mutants to the vacuole.
酵母Kex2蛋白酶(Kex2p)定位于反式高尔基体网络(TGN)需要Kex2p C端胞质尾部的TGN定位信号(TLS)。TLS的突变通过细胞内(不依赖SEC1)途径加速Kex2p向液泡的转运。相反,网格蛋白重链基因CHC1的失活导致Kex2p和其他高尔基体膜蛋白转运到细胞表面。在此,通过检测温度敏感型CHC1等位基因对野生型(WT)和Kex2p TLS突变体形式转运的影响,评估了两种定位缺陷之间的关系。将chc1-ts细胞转移至37℃使网格蛋白失活,导致WT和TLS突变体形式的Kex2p表现相同。在温度转移后30 - 60分钟内,所有形式的Kex2p均出现在质膜上。Kex2p的TLS突变体形式得以稳定,其半衰期增加至野生型Kex2p的半衰期。网格蛋白重链失活后,分泌泡与质膜融合所需的sec1突变阻断了所有形式Kex2p的液泡蛋白酶依赖性降解,这表明液泡蛋白水解降解需要转运至细胞表面。最后,网格蛋白失活后,所有形式的Kex2p部分通过不依赖液泡蛋白酶的途径降解。chc1-ts和sec1-ts均失活后,Kex2仅通过该途径降解。我们得出结论,网格蛋白失活对Kex2p定位的影响独立于Kex2p C端胞质尾部。尽管这些结果既未证明也未排除Kex2 TLS与网格蛋白依赖性结构之间的直接相互作用,但它们确实表明网格蛋白是Kex2p TLS突变体向液泡进行细胞内转运所必需的。