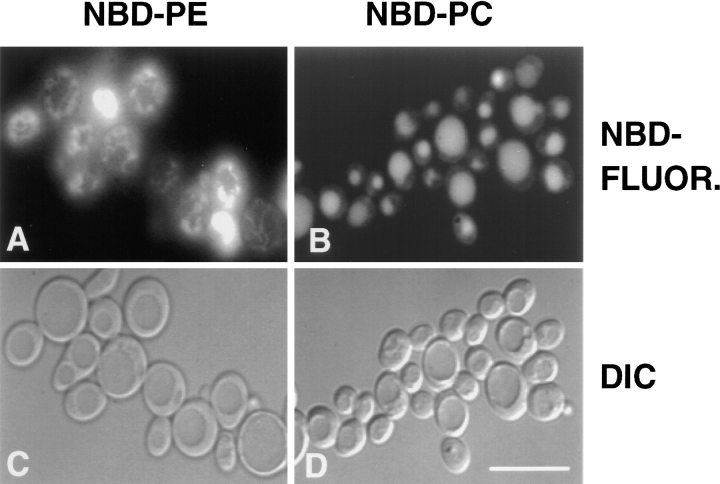
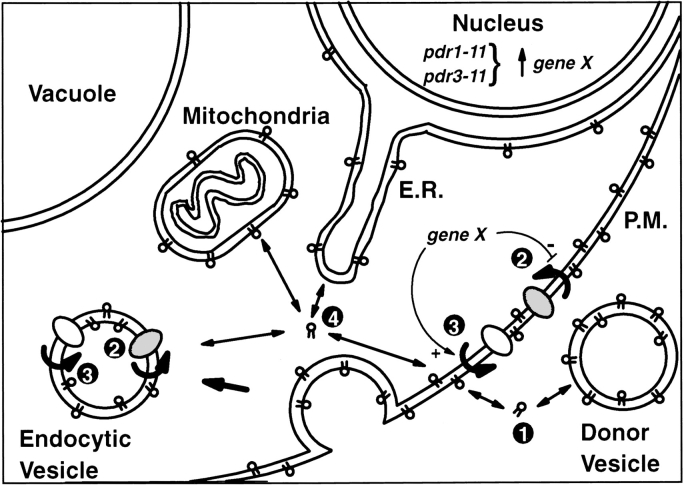
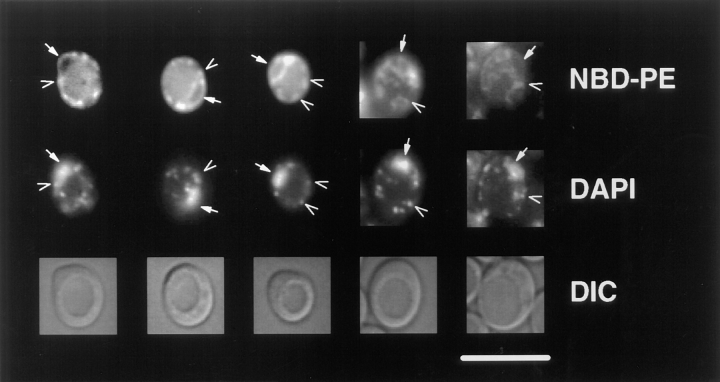
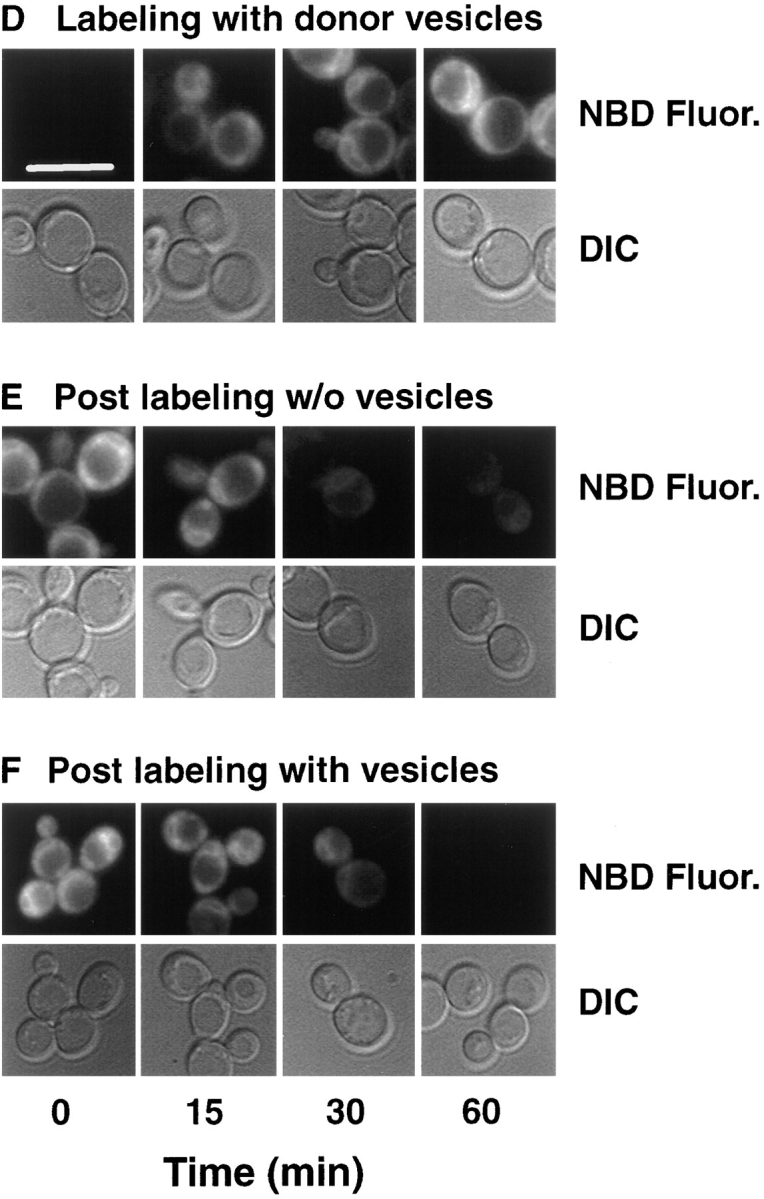
Kean L S, Grant A M, Angeletti C, Mahé Y, Kuchler K, Fuller R S, Nichols J W
Department of Physiology, Emory University School of Medicine, Atlanta, Georgia 30322, USA.
J Cell Biol. 1997 Jul 28;138(2):255-70. doi: 10.1083/jcb.138.2.255.
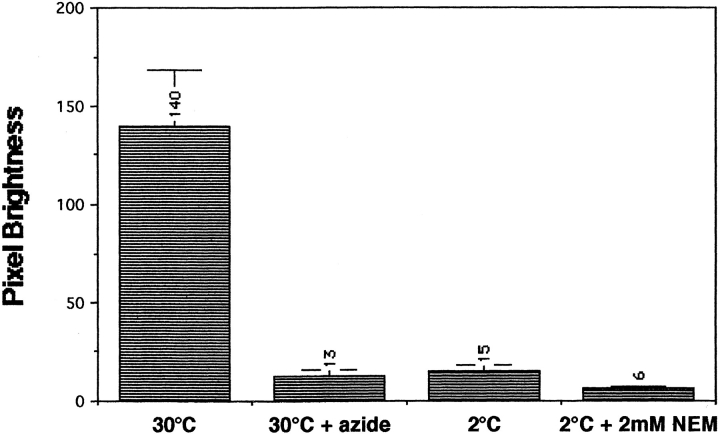
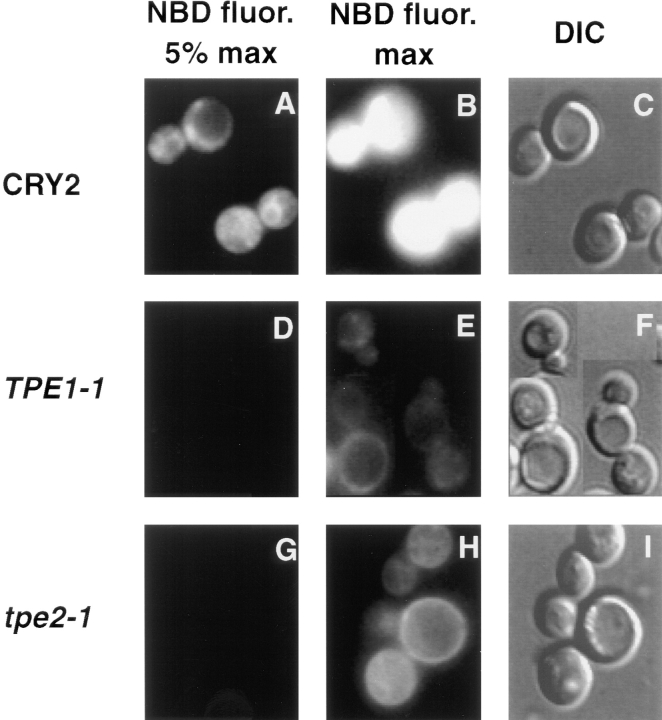
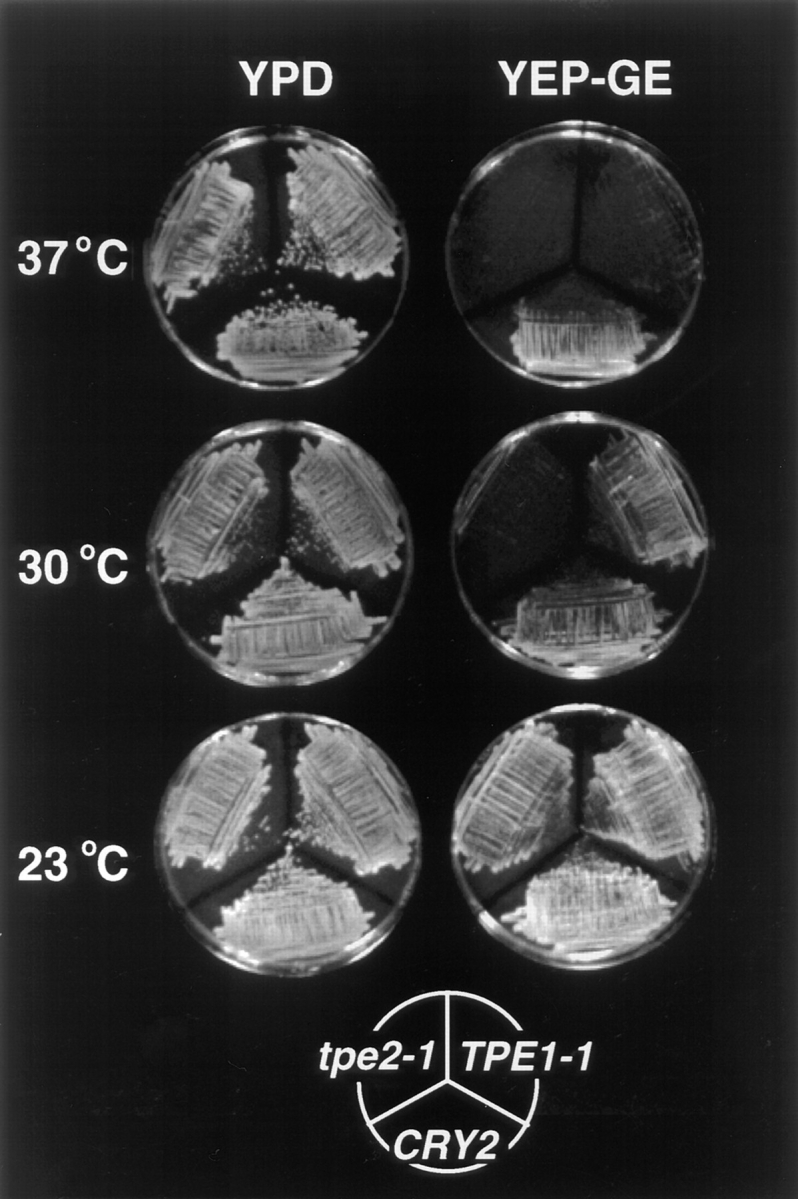
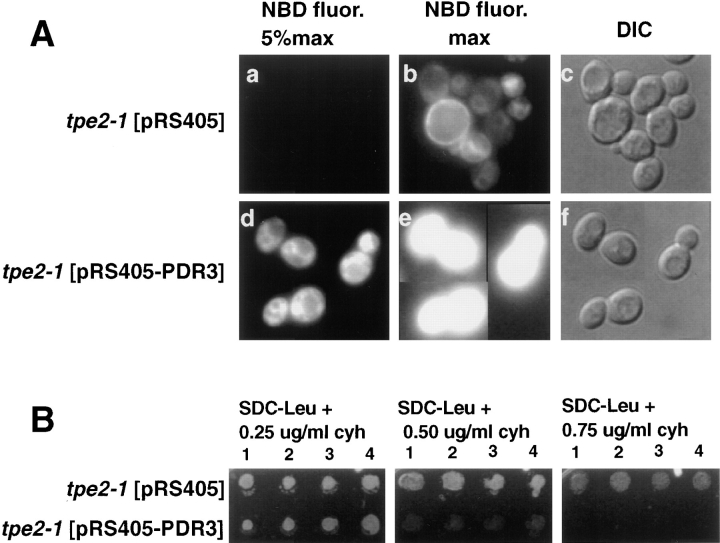
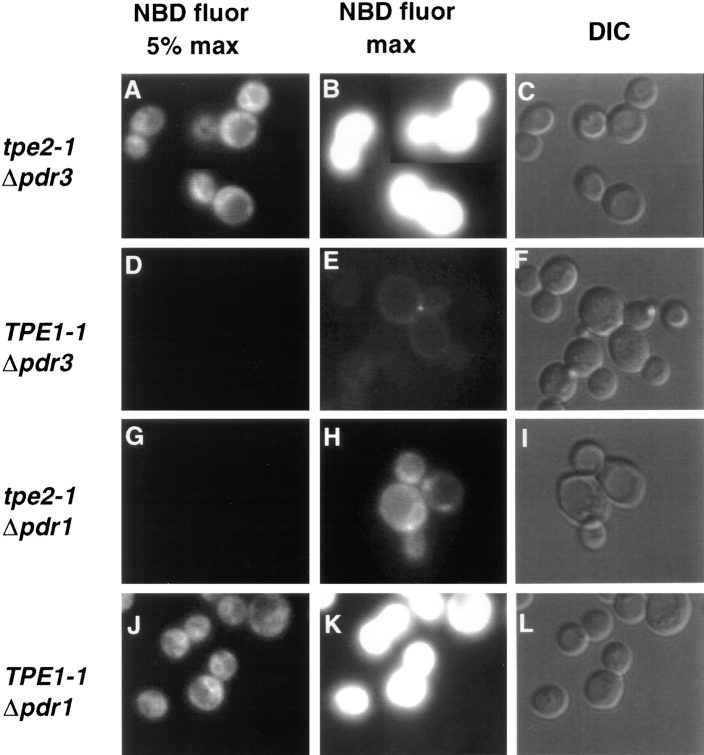
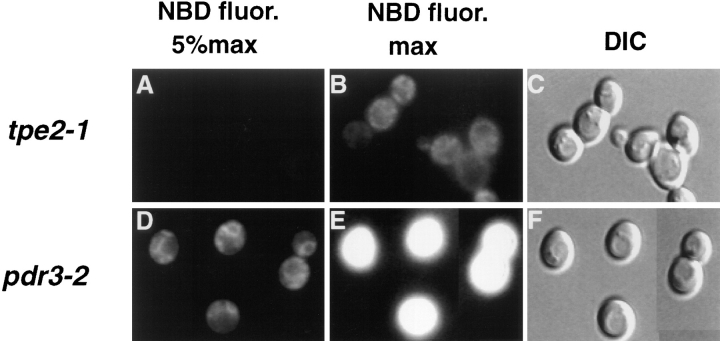
The transcription regulators, PDR1 and PDR3, have been shown to activate the transcription of numerous genes involved in a wide range of functions, including resistance to physical and chemical stress, membrane transport, and organelle function in Saccharomyces cerevisiae. We report here that PDR1 and PDR3 also regulate the transcription of one or more undetermined genes that translocate endogenous and fluorescent-labeled (M-C6-NBD-PE) phosphatidylethanolamine across the plasma membrane. A combination of fluorescence microscopy, fluorometry, and quantitative analysis demonstrated that M-C6-NBD-PE can be translocated both inward and outward across the plasma membrane of yeast cells. Mutants, defective in the accumulation of M-C6-NBD-PE, were isolated by selectively photokilling normal cells that accumulated the fluorescent phospholipid. This led to the isolation of numerous trafficking in phosphatidylethanolamine (tpe) mutants that were defective in intracellular accumulation of M-C6-NBD-PE. Complementation cloning and linkage analysis led to the identification of the dominant mutation TPE1-1 as a new allele of PDR1 and the semidominant mutation tpe2-1 as a new allele of PDR3. The amount of endogenous phosphatidylethanolamine exposed to the outer leaflet of the plasma membrane was measured by covalent labeling with the impermeant amino reagent, trinitrobenzenesulfonic acid. The amount of outer leaflet phosphatidylethanolamine in both mutant strains increased four- to fivefold relative to the parent Tpe+ strain, indicating that the net inward flux of endogenous phosphatidylethanolamine as well as M-C6-NBD-PE was decreased. Targeted deletions of PDR1 in the new allele, PDR1-11, and PDR3 in the new allele, pdr3-11, resulted in normal M-C6-NBD-PE accumulation, confirming that PDR1-11 and pdr3-11 were gain-of-function mutations in PDR1 and PDR3, respectively. Both mutant alleles resulted in resistance to the drugs cycloheximide, oligomycin, and 4-nitroquinoline N-oxide (4-NQO). However, a previously identified drug-resistant allele, pdr3-2, accumulated normal amounts of M-C6-NBD-PE, indicating allele specificity for the loss of M-C6-NBD-PE accumulation. These data demonstrated that PDR1 and PDR3 regulate the net rate of M-C6-NBD-PE translocation (flip-flop) and the steady-state distribution of endogenous phosphatidylethanolamine across the plasma membrane.
转录调节因子PDR1和PDR3已被证明可激活酿酒酵母中许多参与广泛功能的基因的转录,这些功能包括对物理和化学胁迫的抗性、膜运输以及细胞器功能。我们在此报告,PDR1和PDR3还调节一个或多个未确定基因的转录,这些基因可使内源性和荧光标记的(M-C6-NBD-PE)磷脂酰乙醇胺跨质膜转运。荧光显微镜、荧光测定法和定量分析相结合表明,M-C6-NBD-PE可以在酵母细胞质膜内外双向转运。通过选择性光杀死积累荧光磷脂的正常细胞,分离出了在M-C6-NBD-PE积累方面存在缺陷的突变体。这导致分离出许多磷脂酰乙醇胺转运(tpe)突变体,这些突变体在细胞内M-C6-NBD-PE积累方面存在缺陷。互补克隆和连锁分析导致鉴定出显性突变TPE1-1是PDR1的一个新等位基因,半显性突变tpe2-1是PDR3的一个新等位基因。通过用不透膜的氨基试剂三硝基苯磺酸进行共价标记,测量了暴露于质膜外小叶的内源性磷脂酰乙醇胺的量。相对于亲本Tpe+菌株,两个突变菌株中外小叶磷脂酰乙醇胺的量增加了四到五倍,这表明内源性磷脂酰乙醇胺以及M-C6-NBD-PE的净内向通量降低。在新等位基因PDR1-11中靶向缺失PDR1,在新等位基因pdr3-11中靶向缺失PDR3,导致M-C6-NBD-PE正常积累,证实PDR1-11和pdr3-11分别是PDR1和PDR3中的功能获得性突变。这两个突变等位基因均导致对环己酰亚胺、寡霉素和4-硝基喹啉N-氧化物(4-NQO)耐药。然而,先前鉴定的耐药等位基因pdr3-2积累的M-C6-NBD-PE量正常,表明在M-C6-NBD-PE积累缺失方面存在等位基因特异性。这些数据表明,PDR1和PDR3调节M-C6-NBD-PE转运(翻转)的净速率以及内源性磷脂酰乙醇胺在质膜上的稳态分布。