Fitzgerald M L, Wang Z, Park P W, Murphy G, Bernfield M
Division of Newborn Medicine, Children's Hospital, Harvard Medical School, Boston, Massachusetts 02115, USA.
J Cell Biol. 2000 Feb 21;148(4):811-24. doi: 10.1083/jcb.148.4.811.
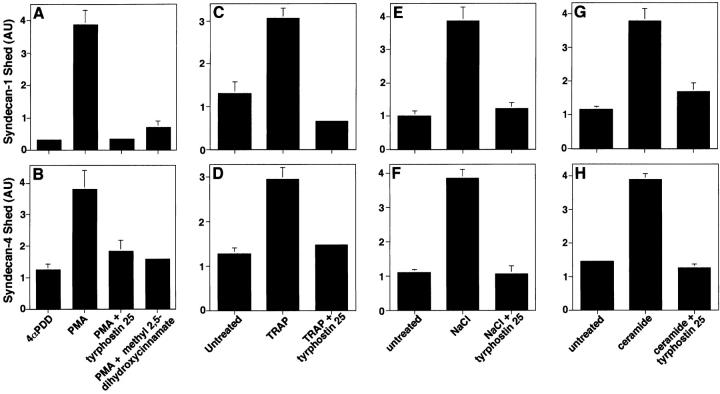
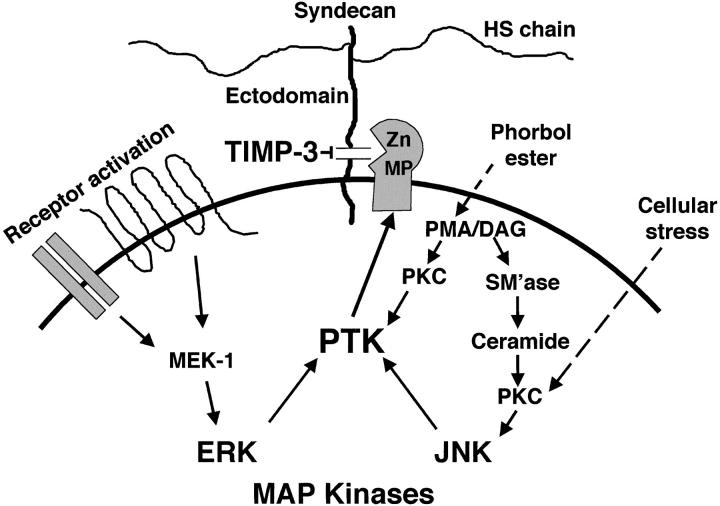
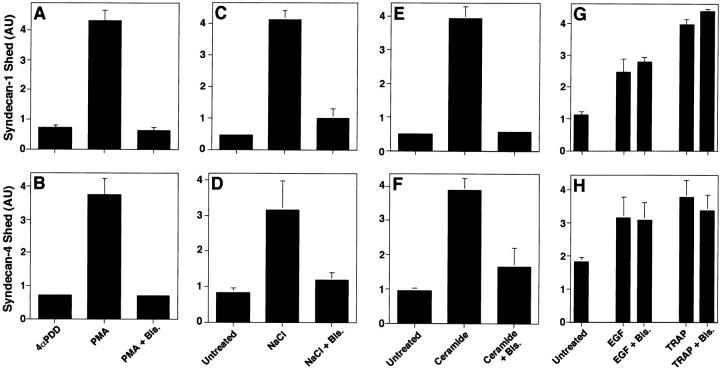
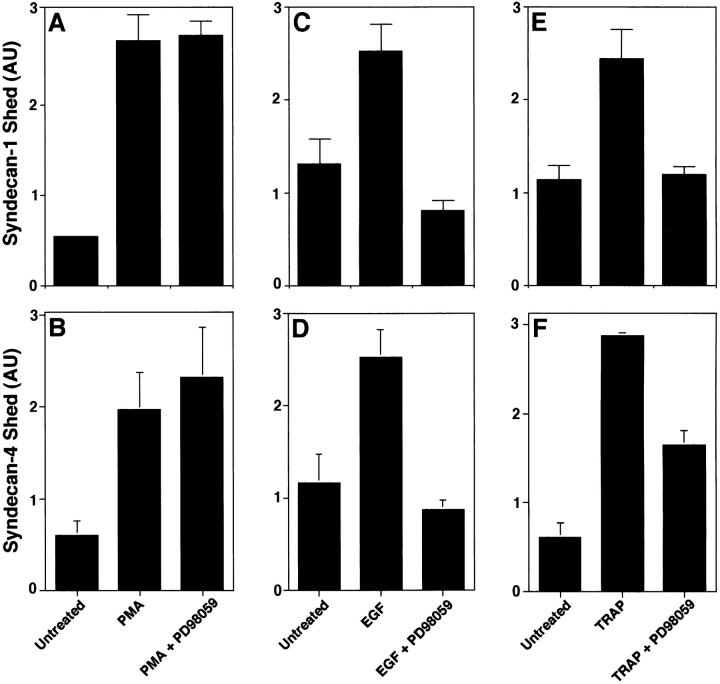
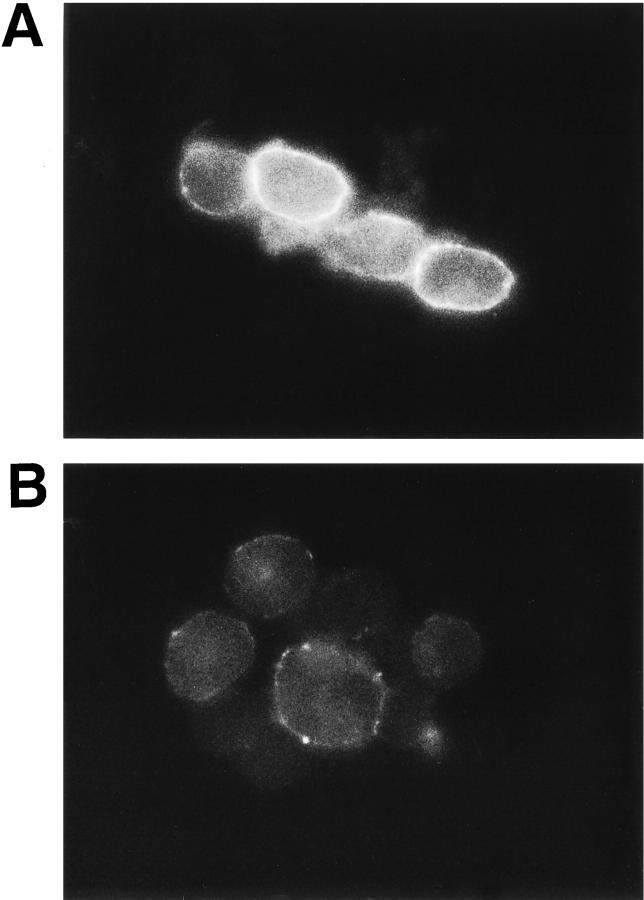
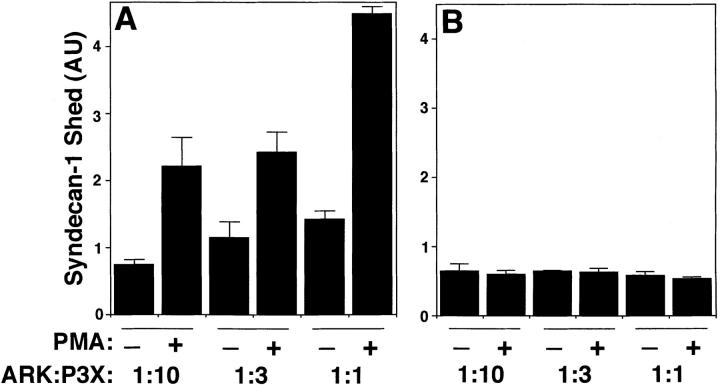
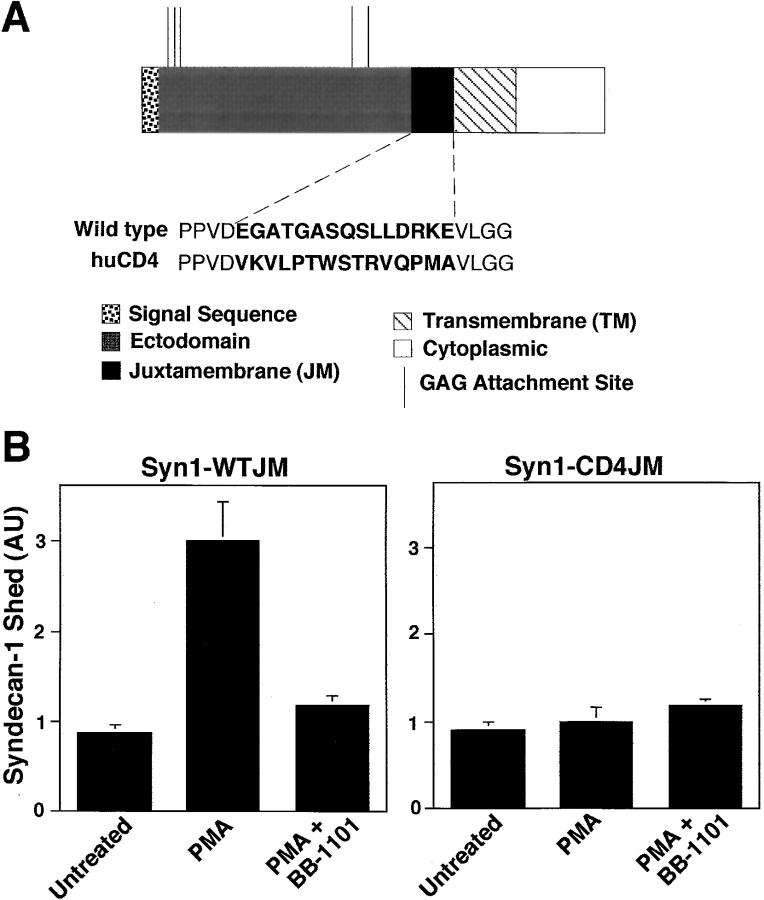
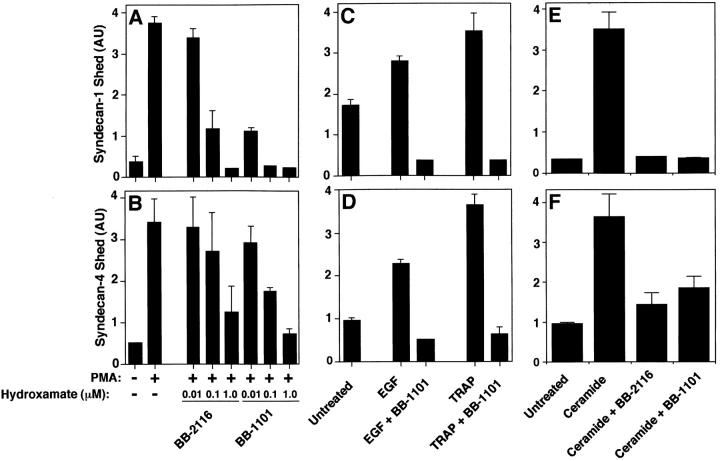
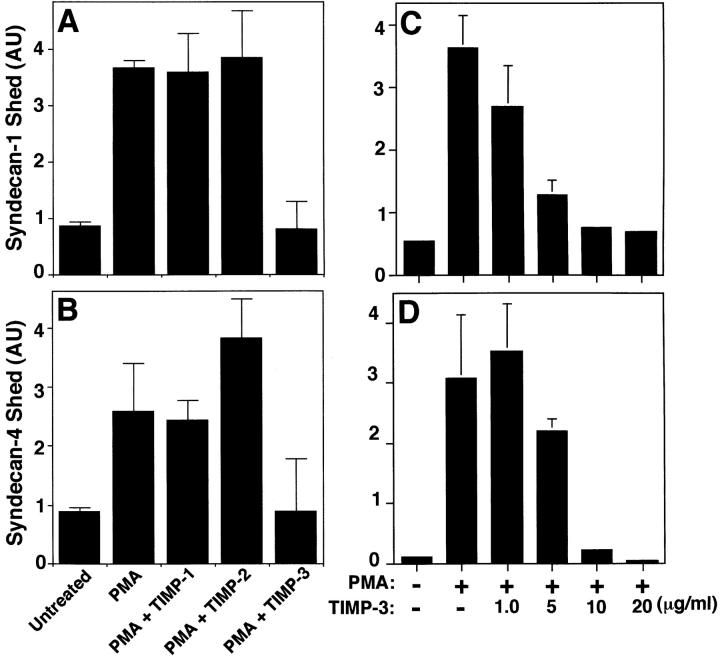
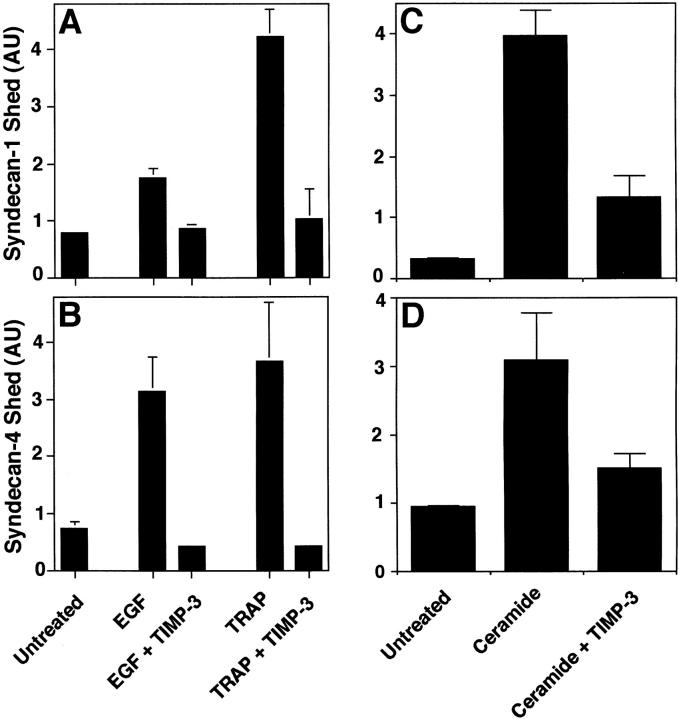
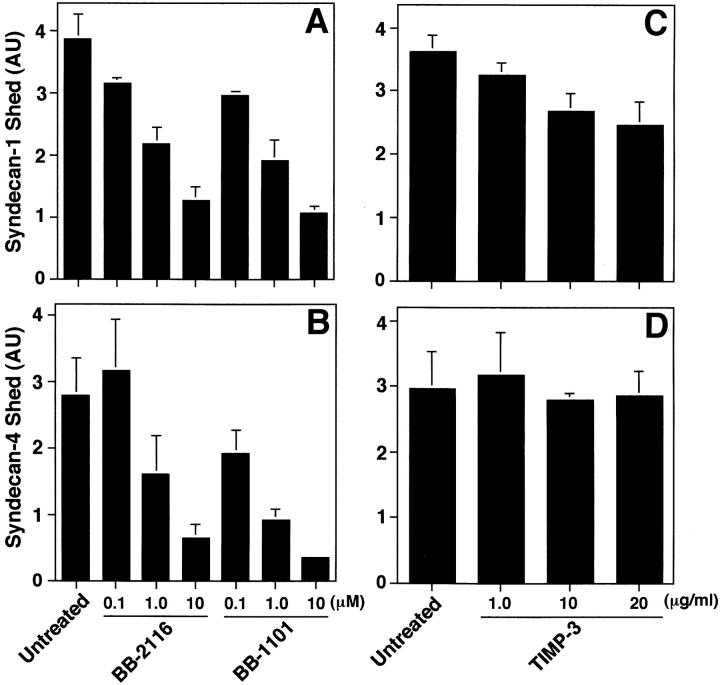
The syndecan family of four transmembrane heparan sulfate proteoglycans binds a variety of soluble and insoluble extracellular effectors. Syndecan extracellular domains (ectodomains) can be shed intact by proteolytic cleavage of their core proteins, yielding soluble proteoglycans that retain the binding properties of their cell surface precursors. Shedding is accelerated by PMA activation of protein kinase C, and by ligand activation of the thrombin (G-protein-coupled) and EGF (protein tyrosine kinase) receptors (Subramanian, S.V., M.L. Fitzgerald, and M. Bernfield. 1997. J. Biol. Chem. 272:14713-14720). Syndecan-1 and -4 ectodomains are found in acute dermal wound fluids, where they regulate growth factor activity (Kato, M., H. Wang, V. Kainulainen, M.L. Fitzgerald, S. Ledbetter, D.M. Ornitz, and M. Bernfield. 1998. Nat. Med. 4:691-697) and proteolytic balance (Kainulainen, V., H. Wang, C. Schick, and M. Bernfield. 1998. J. Biol. Chem. 273:11563-11569). However, little is known about how syndecan ectodomain shedding is regulated. To elucidate the mechanisms that regulate syndecan shedding, we analyzed several features of the process that sheds the syndecan-1 and -4 ectodomains. We find that shedding accelerated by various physiologic agents involves activation of distinct intracellular signaling pathways; and the proteolytic activity responsible for cleavage of syndecan core proteins, which is associated with the cell surface, can act on unstimulated adjacent cells, and is specifically inhibited by TIMP-3, a matrix-associated metalloproteinase inhibitor. In addition, we find that the syndecan-1 core protein is cleaved on the cell surface at a juxtamembrane site; and the proteolytic activity responsible for accelerated shedding differs from that involved in constitutive shedding of the syndecan ectodomains. These results demonstrate the existence of highly regulated mechanisms that can rapidly convert syndecans from cell surface receptors or coreceptors to soluble heparan sulfate proteoglycan effectors. Because the shed ectodomains are found and function in vivo, regulation of syndecan ectodomain shedding by physiological mediators indicates that shedding is a response to specific developmental and pathophysiological cues.
四种跨膜硫酸乙酰肝素蛋白聚糖的syndecan家族可结合多种可溶性和不溶性细胞外效应分子。Syndecan细胞外结构域(胞外域)可通过其核心蛋白的蛋白水解切割而完整脱落,产生可溶性蛋白聚糖,这些蛋白聚糖保留了其细胞表面前体的结合特性。蛋白激酶C的PMA激活、凝血酶(G蛋白偶联)和EGF(蛋白酪氨酸激酶)受体的配体激活可加速脱落(Subramanian, S.V., M.L. Fitzgerald, and M. Bernfield. 1997. J. Biol. Chem. 272:14713 - 14720)。在急性皮肤伤口液中发现了Syndecan - 1和 - 4胞外域,它们在其中调节生长因子活性(Kato, M., H. Wang, V. Kainulainen, M.L. Fitzgerald, S. Ledbetter, D.M. Ornitz, and M. Bernfield. 1998. Nat. Med. 4:691 - 697)和蛋白水解平衡(Kainulainen, V., H. Wang, C. Schick, and M. Bernfield. 1998. J. Biol. Chem. 273:11563 - 11569)。然而,关于syndecan胞外域脱落如何被调节知之甚少。为了阐明调节syndecan脱落的机制,我们分析了syndecan - 1和 - 4胞外域脱落过程的几个特征。我们发现,各种生理因子加速的脱落涉及不同细胞内信号通路的激活;负责切割syndecan核心蛋白的蛋白水解活性与细胞表面相关,可作用于未受刺激的相邻细胞,并被基质相关金属蛋白酶抑制剂TIMP - 3特异性抑制。此外,我们发现syndecan - 1核心蛋白在细胞表面的近膜位点被切割;负责加速脱落的蛋白水解活性与参与syndecan胞外域组成型脱落的活性不同。这些结果证明存在高度调节的机制,可迅速将syndecans从细胞表面受体或共受体转变为可溶性硫酸乙酰肝素蛋白聚糖效应分子。由于脱落的胞外域在体内被发现并发挥功能,生理介质对syndecan胞外域脱落的调节表明脱落是对特定发育和病理生理线索的一种反应。