Colegrove S L, Albrecht M A, Friel D D
Department of Neuroscience, Case Western Reserve University, Cleveland, Ohio 44106, USA.
J Gen Physiol. 2000 Mar;115(3):351-70. doi: 10.1085/jgp.115.3.351.
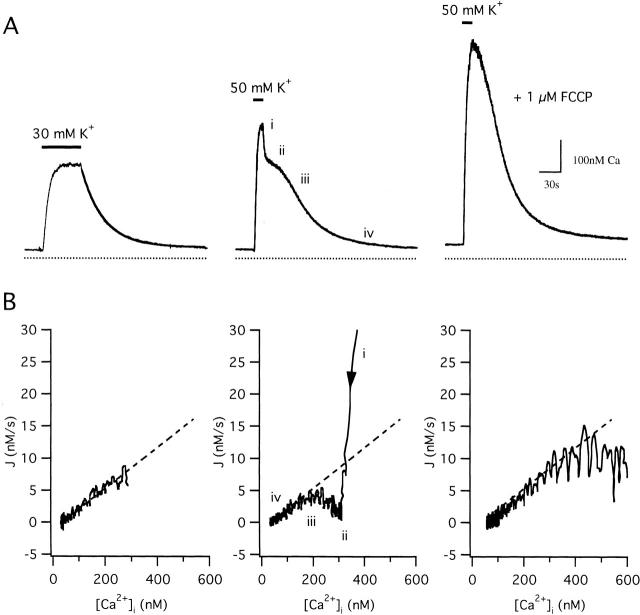
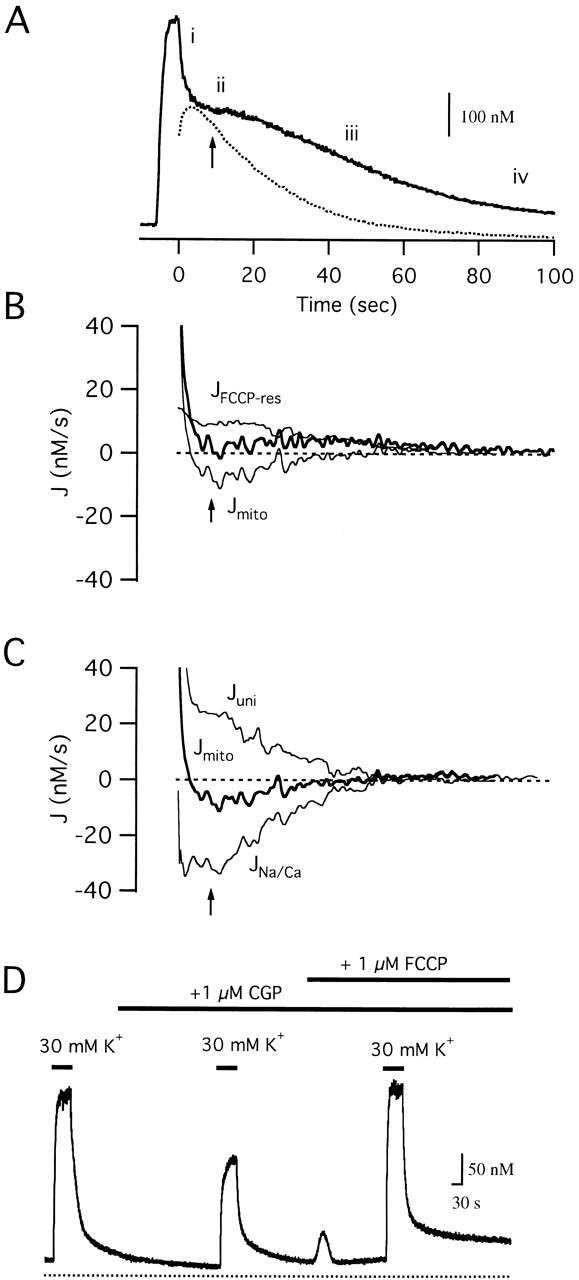
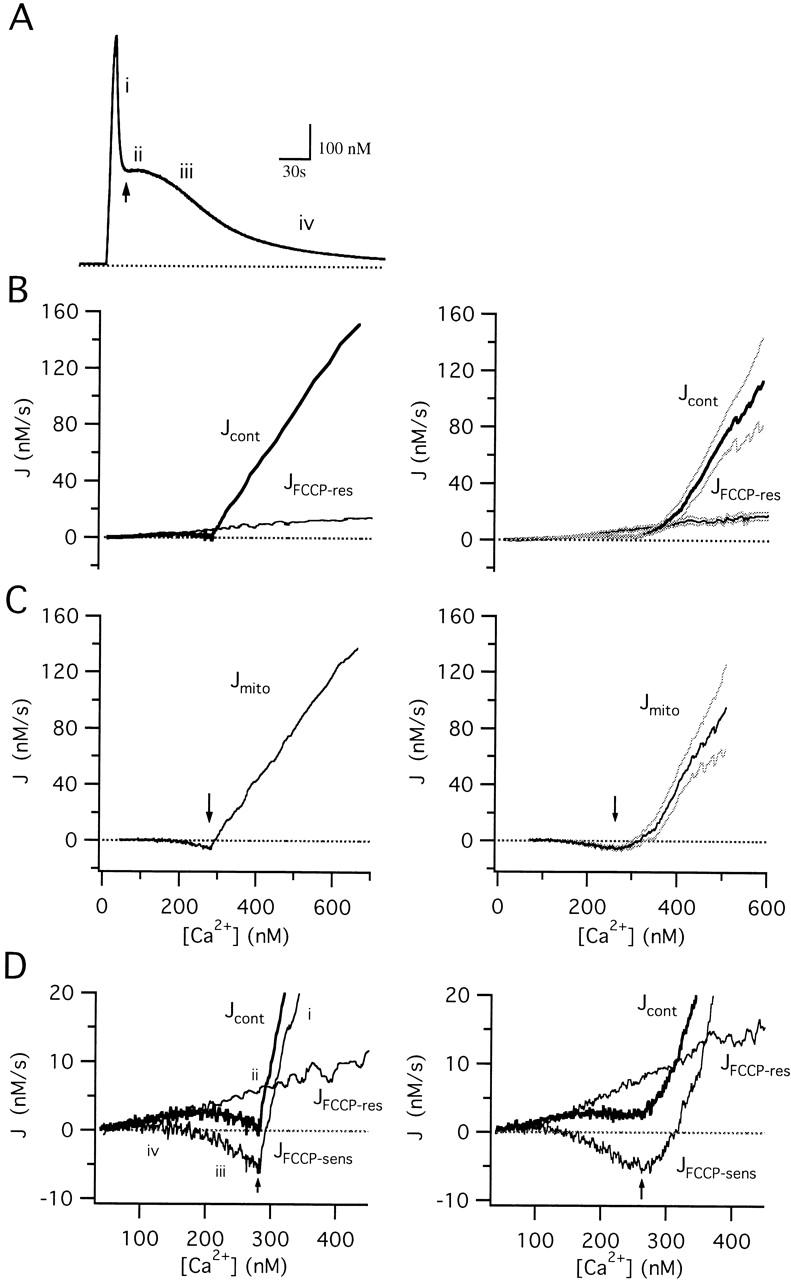
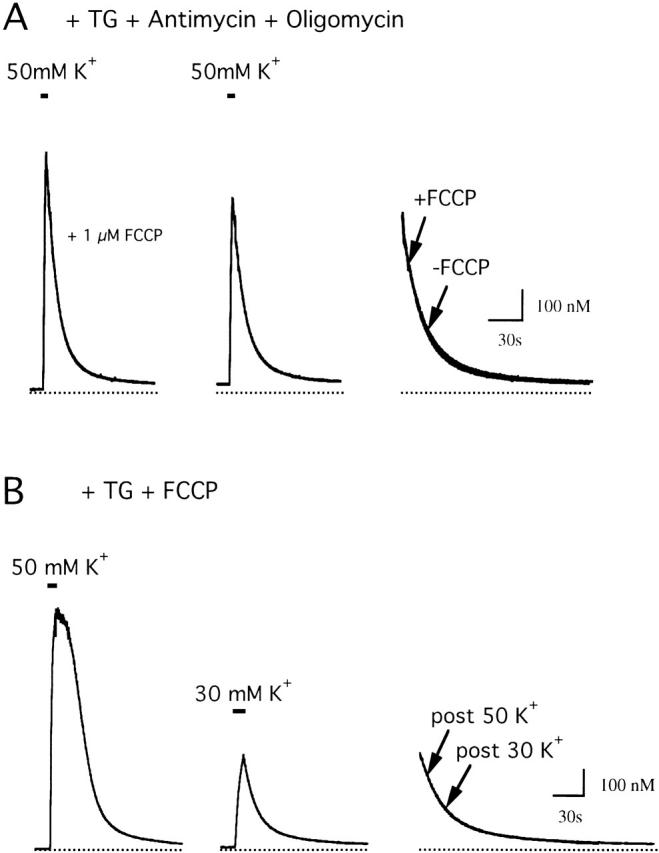
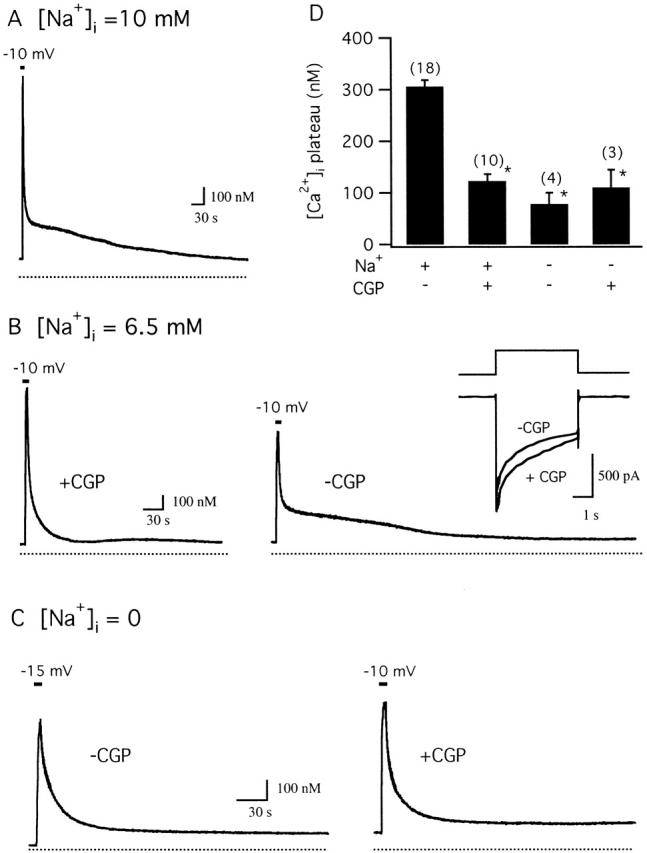
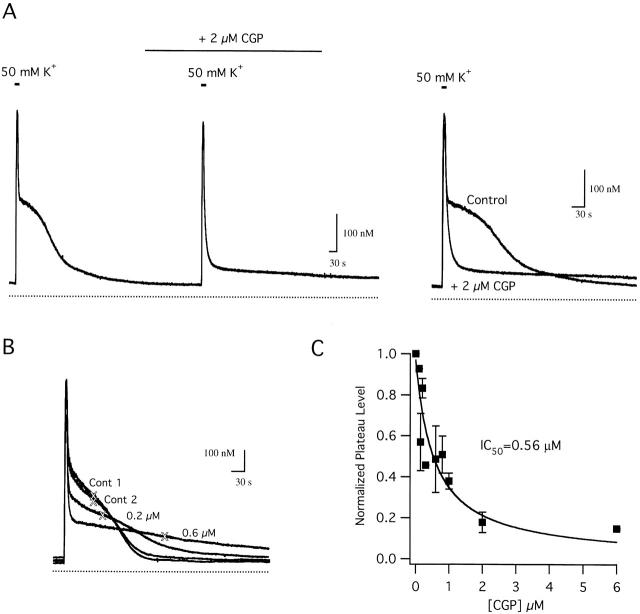
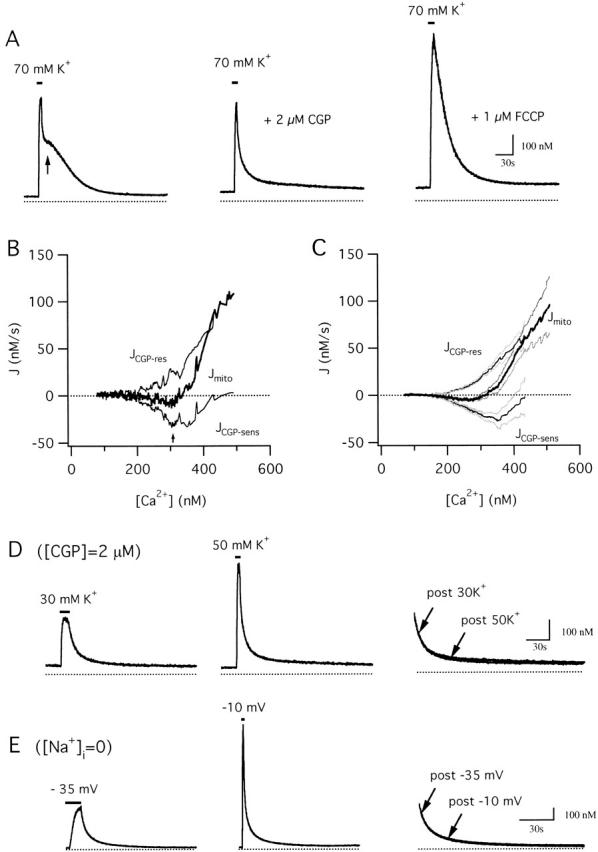
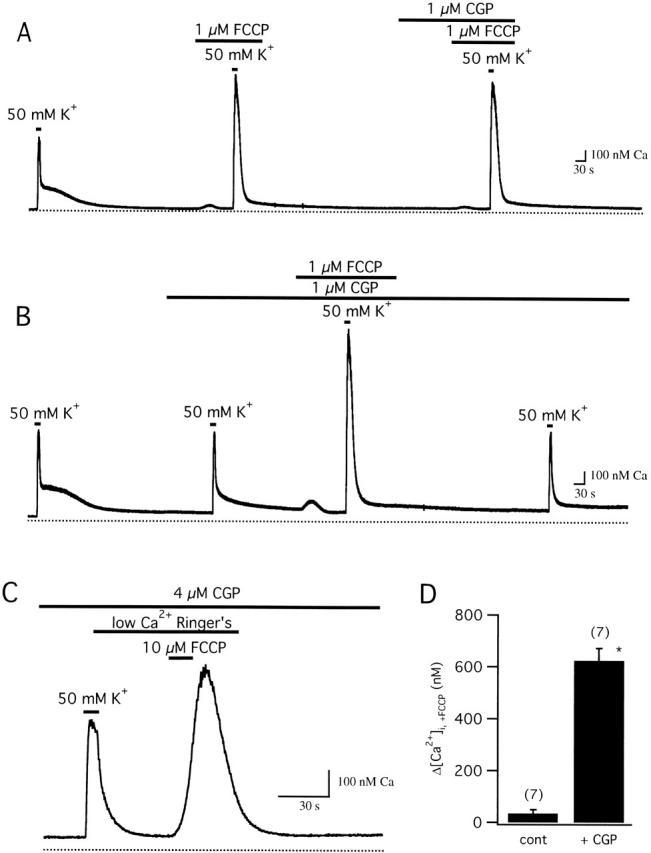
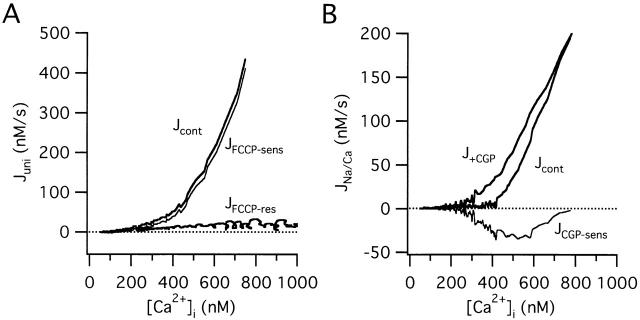
We studied how mitochondrial Ca2+ transport influences Ca2+ dynamics in sympathetic neurons. Cells were treated with thapsigargin to inhibit Ca2+ accumulation by SERCA pumps and depolarized to elevate [Ca2+(i); the recovery that followed repolarization was then examined. The total Ca2+ flux responsible for the Ca2+ recovery was separated into mitochondrial and nonmitochondrial components based on sensitivity to the proton ionophore FCCP, a selective inhibitor of mitochondrial Ca2+ transport in these cells. The nonmitochondrial flux, representing net Ca2+ extrusion across the plasma membrane, has a simple dependence on Ca2+, while the net mitochondrial flux (J(mito)) is biphasic, indicative of Ca+) accumulation during the initial phase of recovery when Ca2+ is high, and net Ca2+ release during later phases of recovery. During each phase, mitochondrial Ca2+ transport has distinct effects on recovery kinetics. J(mito) was separated into components representing mitochondrial Ca2+ uptake and release based on sensitivity to the specific mitochondrial Na(+)/Ca2+ exchange inhibitor, CGP 37157 (CGP). The CGP-resistant (uptake) component of J(mito) increases steeply with Ca2+, as expected for transport by the mitochondrial uniporter. The CGP-sensitive (release) component is inhibited by lowering the intracellular Na(+) concentration and depends on both intra- and extramitochondrial Ca2+ concentration, as expected for the Na(+)/Ca2+ exchanger. Above approximately 400 nM Ca2+, net mitochondrial Ca2+ transport is dominated by uptake and is largely insensitive to CGP. When Ca2+ is approximately 200-300 nM, the net mitochondrial flux is small but represents the sum of much larger uptake and release fluxes that largely cancel. Thus, mitochondrial Ca2+ transport occurs in situ at much lower concentrations than previously thought, and may provide a mechanism for quantitative control of ATP production after brief or low frequency stimuli that raise Ca(2+) to levels below approximately 500 nM.
我们研究了线粒体Ca2+转运如何影响交感神经元中的Ca2+动态变化。用毒胡萝卜素处理细胞以抑制SERCA泵的Ca2+积累,并使其去极化以提高[Ca2+(i)];然后检查复极化后的恢复情况。基于对质子离子载体FCCP(这些细胞中线粒体Ca2+转运的选择性抑制剂)的敏感性,将负责Ca2+恢复的总Ca2+通量分为线粒体和非线粒体成分。非线粒体通量代表跨质膜的净Ca2+外流,它对Ca2+有简单的依赖性,而线粒体净通量(J(mito))是双相的,表明在恢复初期当Ca2+较高时Ca+积累,以及在恢复后期净Ca2+释放。在每个阶段,线粒体Ca2+转运对恢复动力学有不同的影响。基于对特异性线粒体Na(+)/Ca2+交换抑制剂CGP 37157(CGP)的敏感性,J(mito)被分为代表线粒体Ca2+摄取和释放的成分。正如线粒体单向转运体转运所预期的那样,J(mito)的CGP抗性(摄取)成分随Ca2+急剧增加。正如Na(+)/Ca2+交换体所预期的那样,CGP敏感(释放)成分通过降低细胞内Na(+)浓度而受到抑制,并且依赖于线粒体内外的Ca2+浓度。当Ca2+高于约400 nM时,线粒体净Ca2+转运以摄取为主,并且对CGP基本不敏感。当Ca2+约为200 - 300 nM时,线粒体净通量较小,但代表了在很大程度上相互抵消的更大的摄取和释放通量之和。因此,线粒体Ca2+转运在比以前认为的低得多的浓度下原位发生,并且可能为短暂或低频刺激将Ca(2+)提高到低于约500 nM的水平后ATP产生的定量控制提供一种机制。