Schweizer Liang, Varmus Harold
Cell Biology Program, Sloan-Kettering Institute for Cancer Research, 1275 York Avenue, New York, NY 10021, USA.
BMC Cell Biol. 2003 May 2;4:4. doi: 10.1186/1471-2121-4-4.
Wnt/Wingless (Wg) signals are transduced by seven-transmembrane Frizzleds (Fzs) and the single-transmembrane LDL-receptor-related proteins 5 or 6 (LRP5/6) or Arrow. The aminotermini of LRP and Fz were reported to associate only in the presence of Wnt, implying that Wnt ligands form a trimeric complex with two different receptors. However, it was recently reported that LRPs activate the Wnt/beta-catenin pathway by binding to Axin in a Dishevelled--independent manner, while Fzs transduce Wnt signals through Dishevelled to stabilize beta-catenin. Thus, it is possible that Wnt proteins form separate complexes with Fzs and LRPs, transducing Wnt signals separately, but converging downstream in the Wnt/beta-catenin pathway. The question then arises whether both receptors are absolutely required to transduce Wnt signals.
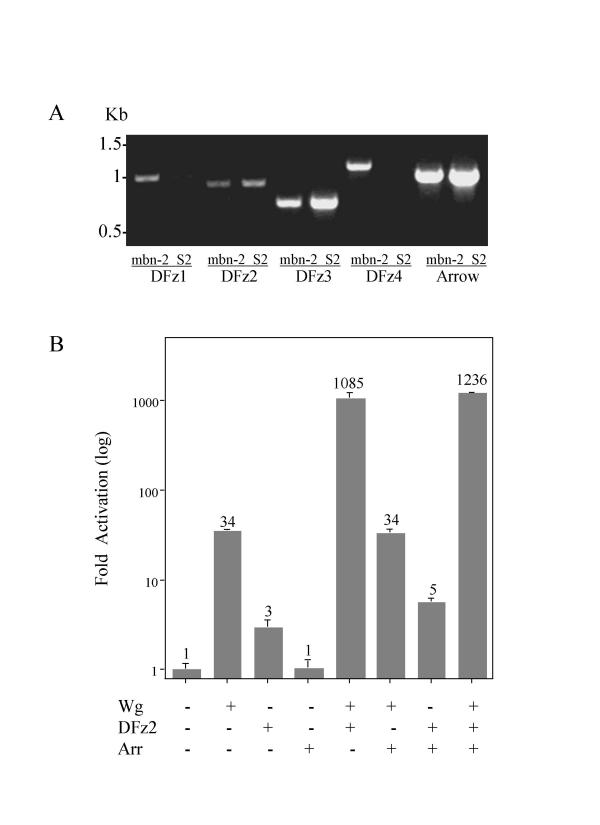
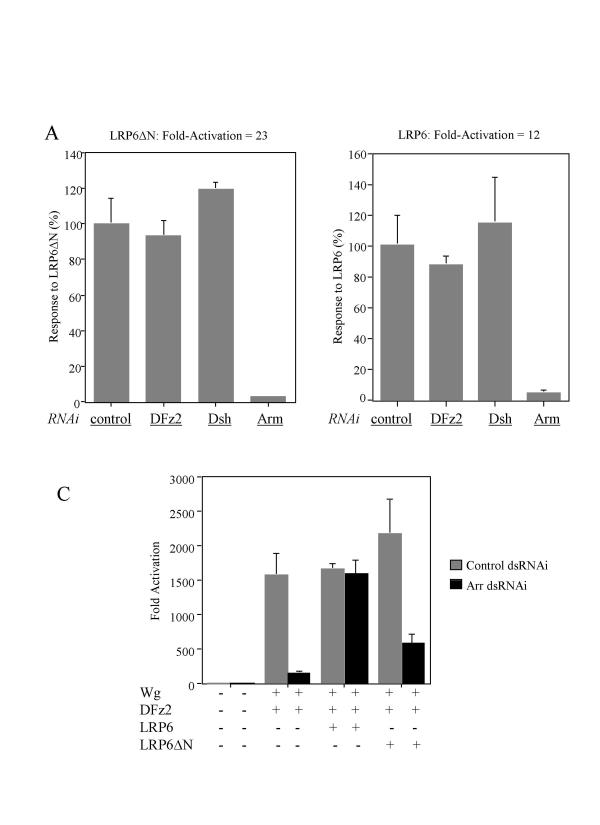
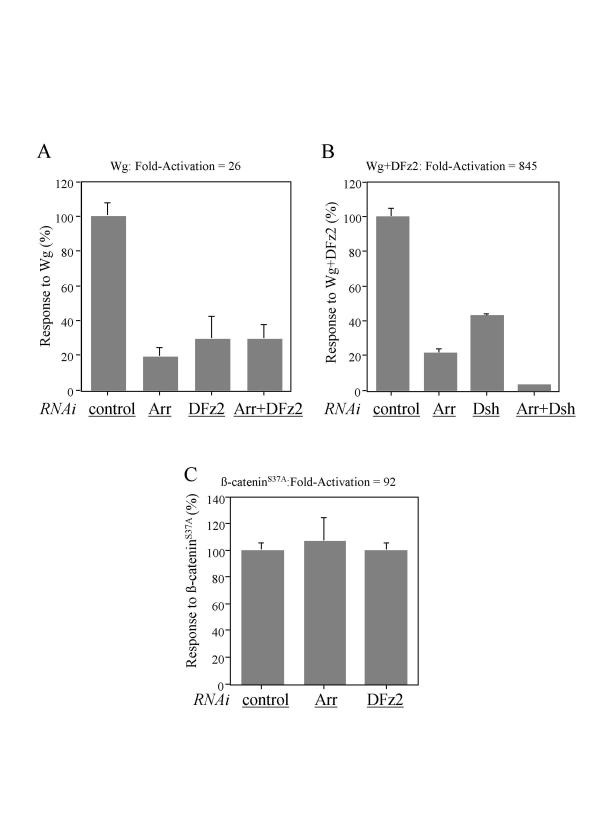
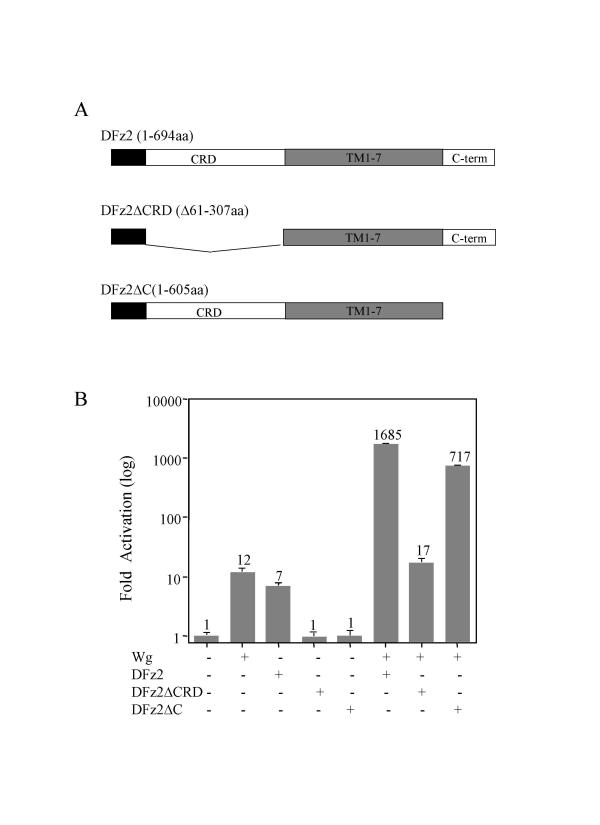
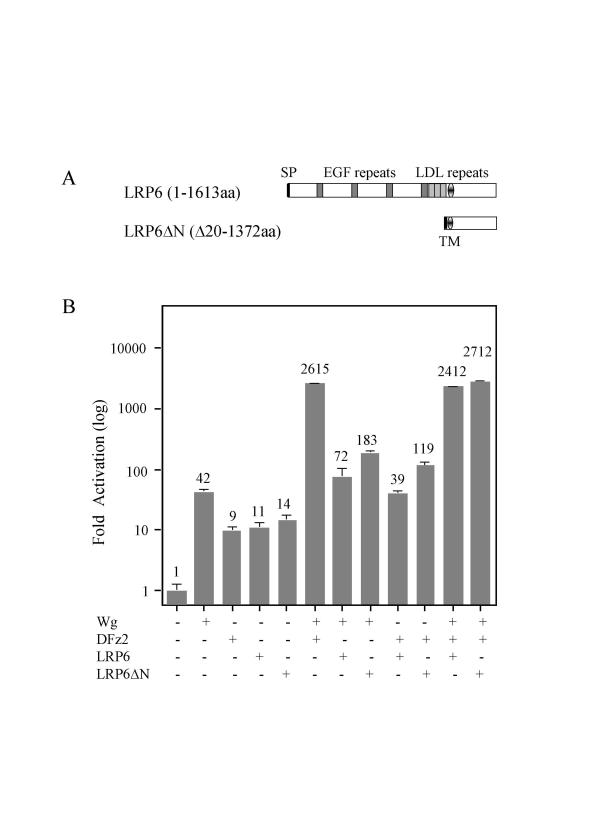
We have established a sensitive luciferase reporter assay in Drosophila S2 cells to determine the level of Wg--stimulated signaling. We demonstrate here that Wg can synergize with DFz2 and function cooperatively with LRP to activate the beta-catenin/Armadillo signaling pathway. Double-strand RNA interference that disrupts the synthesis of either receptor type dramatically impairs Wg signaling activity. Importantly, the pronounced synergistic effect of adding Wg and DFz2 is dependent on Arrow and Dishevelled. The synergy requires the cysteine-rich extracellular domain of DFz2, but not its carboxyterminus. Finally, mammalian LRP6 and its activated forms, which lack most of the extracellular domain of the protein, can activate the Wg signaling pathway and cooperate with Wg and DFz2 in S2 cells. We also show that the aminoterminus of LRP/Arr is required for the synergy between Wg and DFz2.
Our study indicates that Wg signal transduction in S2 cells depends on the function of both LRPs and DFz2, and the results are consistent with the proposal that Wnt/Wg signals through the aminoterminal domains of its dual receptors, activating target genes through Dishevelled.
Wnt/Wingless(Wg)信号由七次跨膜的卷曲蛋白(Fzs)以及单次跨膜的低密度脂蛋白受体相关蛋白5或6(LRP5/6)或箭蛋白(Arrow)进行转导。据报道,LRP和Fz的氨基末端仅在Wnt存在时才会结合,这意味着Wnt配体与两种不同的受体形成三聚体复合物。然而,最近有报道称,LRP通过以一种不依赖于散乱蛋白(Dishevelled)的方式与轴蛋白(Axin)结合来激活Wnt/β-连环蛋白信号通路,而Fzs则通过Dishevelled转导Wnt信号以稳定β-连环蛋白。因此,Wnt蛋白有可能分别与Fzs和LRPs形成复合物,分别转导Wnt信号,但在Wnt/β-连环蛋白信号通路的下游汇聚。那么问题就出现了,两种受体对于转导Wnt信号是否都是绝对必需的。
我们在果蝇S2细胞中建立了一种灵敏的荧光素酶报告基因检测方法,以确定Wg刺激的信号水平。我们在此证明,Wg可以与DFz2协同作用,并与LRP合作激活β-连环蛋白/犰狳蛋白(Armadillo)信号通路。破坏任一受体类型合成的双链RNA干扰会显著损害Wg信号活性。重要的是,添加Wg和DFz2所产生的明显协同效应依赖于箭蛋白和Dishevelled。这种协同作用需要DFz2富含半胱氨酸的细胞外结构域,但不需要其羧基末端。最后,哺乳动物的LRP6及其激活形式(其缺乏该蛋白的大部分细胞外结构域)可以激活Wg信号通路,并在S2细胞中与Wg和DFz2协同作用。我们还表明,LRP/箭蛋白的氨基末端是Wg与DFz2之间协同作用所必需的。
我们的研究表明,S2细胞中的Wg信号转导依赖于LRP和DFz2的功能,并且结果与Wnt/Wg通过其双受体的氨基末端结构域进行信号转导、通过Dishevelled激活靶基因的提议一致。