Basso Claudia, Vergani Paola, Nairn Angus C, Gadsby David C
Laboratory of Cardiac/Membrane Physiology, The Rockefeller University, 1230 York Ave., New York, NY 10021, USA.
J Gen Physiol. 2003 Sep;122(3):333-48. doi: 10.1085/jgp.200308798.
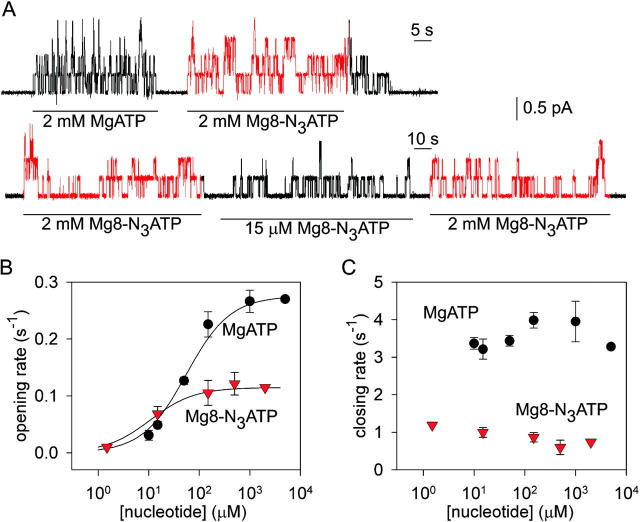
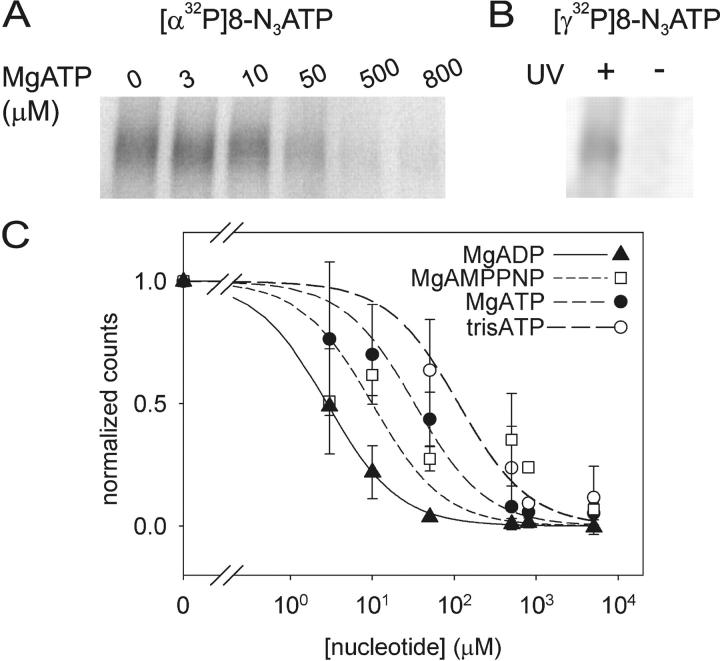
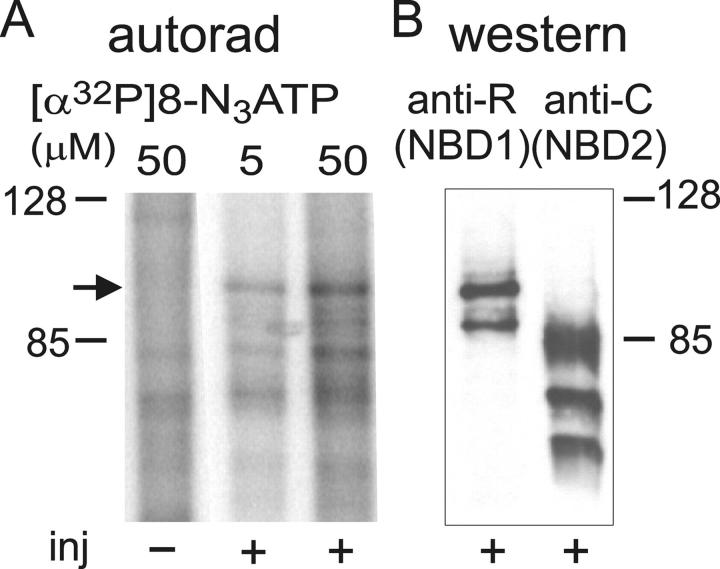
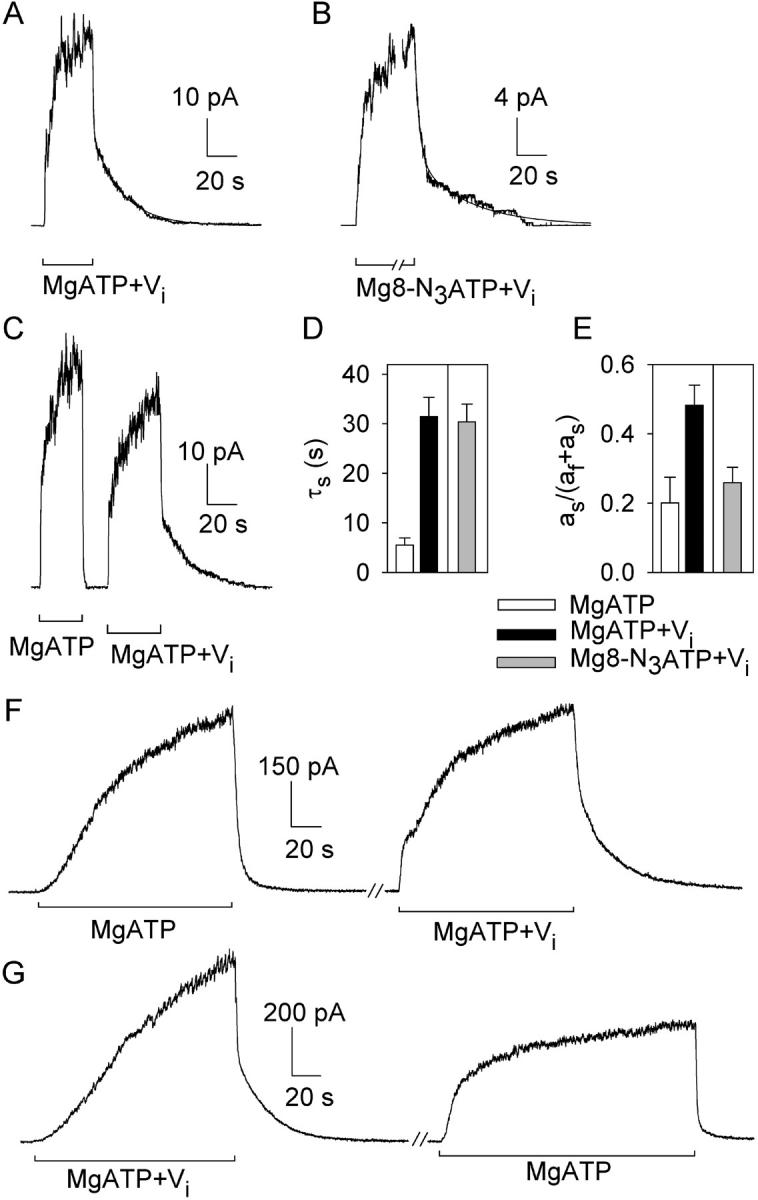
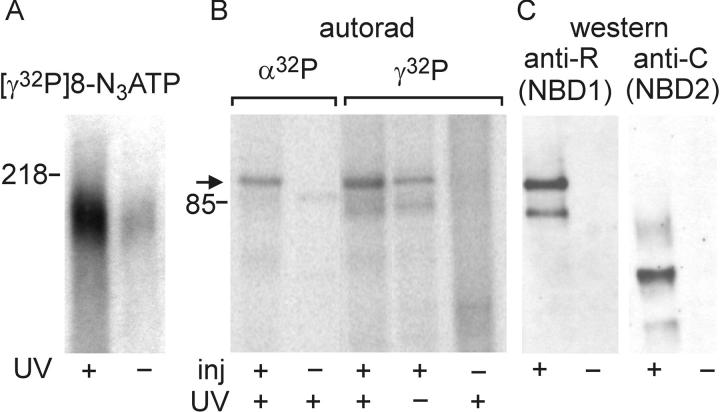
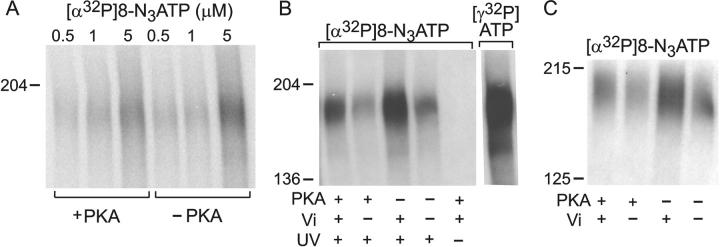
CFTR, the protein defective in cystic fibrosis, functions as a Cl- channel regulated by cAMP-dependent protein kinase (PKA). CFTR is also an ATPase, comprising two nucleotide-binding domains (NBDs) thought to bind and hydrolyze ATP. In hydrolyzable nucleoside triphosphates, PKA-phosphorylated CFTR channels open into bursts, lasting on the order of a second, from closed (interburst) intervals of a second or more. To investigate nucleotide interactions underlying channel gating, we examined photolabeling by [alpha32P]8-N3ATP or [gamma32P]8-N3ATP of intact CFTR channels expressed in HEK293T cells or Xenopus oocytes. We also exploited split CFTR channels to distinguish photolabeling at NBD1 from that at NBD2. To examine simple binding of nucleotide in the absence of hydrolysis and gating reactions, we photolabeled after incubation at 0 degrees C with no washing. Nucleotide interactions under gating conditions were probed by photolabeling after incubation at 30 degrees C, with extensive washing, also at 30 degrees C. Phosphorylation of CFTR by PKA only slightly influenced photolabeling after either protocol. Strikingly, at 30 degrees C nucleotide remained tightly bound at NBD1 for many minutes, in the form of nonhydrolyzed nucleoside triphosphate. As nucleotide-dependent gating of CFTR channels occurred on the time scale of seconds under comparable conditions, this suggests that the nucleotide interactions, including hydrolysis, that time CFTR channel opening and closing occur predominantly at NBD2. Vanadate also appeared to act at NBD2, presumably interrupting its hydrolytic cycle, and markedly delayed termination of channel open bursts. Vanadate somewhat increased the magnitude, but did not alter the rate, of the slow loss of nucleotide tightly bound at NBD1. Kinetic analysis of channel gating in Mg8-N3ATP or MgATP reveals that the rate-limiting step for CFTR channel opening at saturating [nucleotide] follows nucleotide binding to both NBDs. We propose that ATP remains tightly bound or occluded at CFTR's NBD1 for long periods, that binding of ATP at NBD2 leads to channel opening wherupon its hydrolysis prompts channel closing, and that phosphorylation acts like an automobile clutch that engages the NBD events to drive gating of the transmembrane ion pore.
囊性纤维化中存在缺陷的蛋白质CFTR,作为一种受环磷酸腺苷依赖性蛋白激酶(PKA)调节的氯离子通道发挥作用。CFTR也是一种ATP酶,由两个被认为可结合并水解ATP的核苷酸结合结构域(NBD)组成。在可水解的核苷三磷酸中,PKA磷酸化的CFTR通道会从一秒或更长时间的关闭(爆发间期)状态突然打开,持续约一秒钟。为了研究通道门控背后的核苷酸相互作用,我们检测了在HEK293T细胞或非洲爪蟾卵母细胞中表达的完整CFTR通道被[α32P]8-N3ATP或[γ32P]8-N3ATP光标记的情况。我们还利用拆分的CFTR通道来区分NBD1和NBD2处的光标记。为了检测在没有水解和门控反应的情况下核苷酸的简单结合,我们在0℃孵育后进行光标记且不洗涤。通过在30℃孵育后进行光标记来探测门控条件下的核苷酸相互作用,同样在30℃进行大量洗涤。无论采用哪种方案,PKA对CFTR的磷酸化仅对光标记有轻微影响。引人注目的是,在30℃时,核苷酸以未水解的核苷三磷酸形式在NBD1处紧密结合长达数分钟。由于在类似条件下CFTR通道的核苷酸依赖性门控发生在秒的时间尺度上,这表明决定CFTR通道打开和关闭时间的核苷酸相互作用,包括水解,主要发生在NBD2。钒酸盐似乎也作用于NBD2,大概是中断了其水解循环,并显著延迟了通道开放爆发的终止。钒酸盐在一定程度上增加了紧密结合在NBD1处的核苷酸缓慢丢失的幅度,但没有改变其速率。对Mg8-N3ATP或MgATP中通道门控的动力学分析表明,在饱和[核苷酸]浓度下CFTR通道打开的限速步骤是核苷酸与两个NBD的结合。我们提出,ATP在CFTR的NBD1处长时间紧密结合或被封闭,ATP在NBD2处的结合导致通道打开,随后其水解促使通道关闭,并且磷酸化的作用就像汽车离合器,将NBD事件连接起来以驱动跨膜离子孔的门控。