Stuhlmann H, Berg P
Department of Biochemistry, Beckman Center, Stanford University School of Medicine, California 94305-5307.
J Virol. 1992 Apr;66(4):2378-88. doi: 10.1128/JVI.66.4.2378-2388.1992.
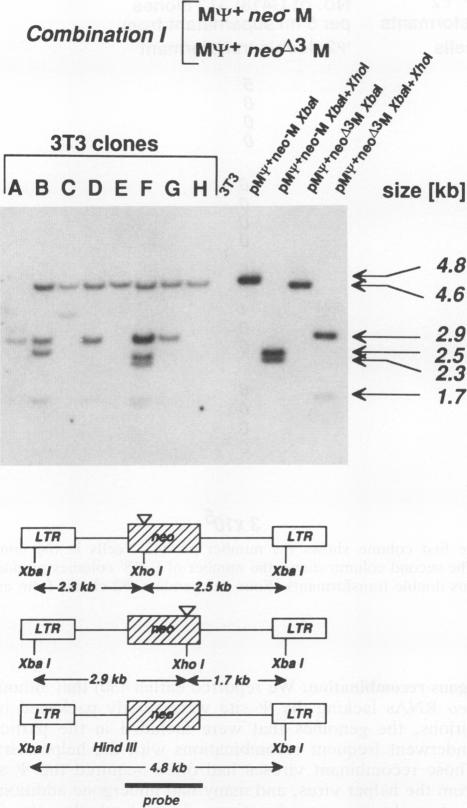
According to prevailing models, the high frequency of recombination in retroviruses occurs during reverse transcription of two genetically different genomes copackaged into virion particles. This view has been tested in our studies of the mechanism of recombination within homologous sequences of two retroviral genomes during a single round of virus replication and in the absence of helper virus. The recombination substrates were Moloney murine leukemia virus-based vectors, each of which contains an altered defective neomycin gene (neo) under the transcriptional control of the 5' long terminal repeat; the 3' sequences of each construct contain either the Moloney murine leukemia virus or simian virus 40 large-T polyadenylation sequence. One neo gene contained a linker insertion mutation at the 5' end (neo minus), and the other contained a deletion and linker insertion at the 3' end (neo delta 3). Each of the mutant neo constructs was introduced into the packaging helper cell line psi 2 by sequential cotransfection, and individual psi 2 double transformants were selected. Supernatant fluids from the cloned psi 2 double transformants were used to infect NIH 3T3 cells, and recombinant neo+ proviruses were detected by their ability to confer G418 resistance during infection of NIH 3T3 cells. Our results show that (i) recombination between a homologous sequence of about 560 bp occurred with a frequency of about 10(-4) per virus replication cycle; (ii) recombination occurred only after the viral RNAs had been packaged into particles, i.e., recombination between the two vector DNAs or between viral RNAs prior to packaging was not detected; and (iii) copackaging of two different genomic RNAs as a heterodimer is a prerequisite for recombination. Furthermore, our results indicate that recombination can occur during the DNA negative-strand synthesis of reverse transcription.
根据流行的模型,逆转录病毒中高频重组发生在共包装进病毒粒子的两个基因不同的基因组的逆转录过程中。在我们关于两轮病毒复制过程中两个逆转录病毒基因组同源序列间重组机制的研究中,且在无辅助病毒的情况下,这一观点得到了验证。重组底物是以莫洛尼鼠白血病病毒为基础的载体,每个载体在5'长末端重复序列的转录控制下含有一个改变的缺陷新霉素基因(neo);每个构建体的3'序列含有莫洛尼鼠白血病病毒或猴病毒40大T抗原聚腺苷酸化序列。一个neo基因在5'端含有一个接头插入突变(neo minus),另一个在3'端含有一个缺失和接头插入(neo delta 3)。通过连续共转染将每个突变的neo构建体导入包装辅助细胞系psi 2,并筛选出单个psi 2双转化体。来自克隆的psi 2双转化体的上清液用于感染NIH 3T3细胞,通过它们在感染NIH 3T3细胞期间赋予G418抗性的能力来检测重组的neo+前病毒。我们的结果表明:(i)约560 bp的同源序列间的重组频率约为每个病毒复制周期10^(-4);(ii)重组仅在病毒RNA被包装进颗粒后发生,即未检测到两个载体DNA之间或包装前病毒RNA之间的重组;(iii)两个不同基因组RNA作为异二聚体共包装是重组的先决条件。此外,我们的结果表明重组可发生在逆转录的DNA负链合成过程中。