Duval N, Krejci E, Grassi J, Coussen F, Massoulié J, Bon S
Laboratoire de Neurobiologie, CNRS URA 295, Ecole Normale Supérieure, France.
EMBO J. 1992 Sep;11(9):3255-61. doi: 10.1002/j.1460-2075.1992.tb05403.x.
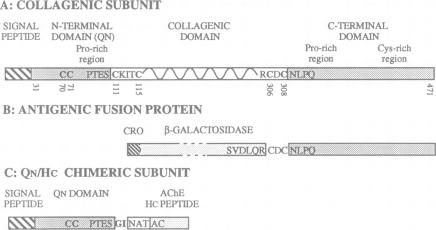
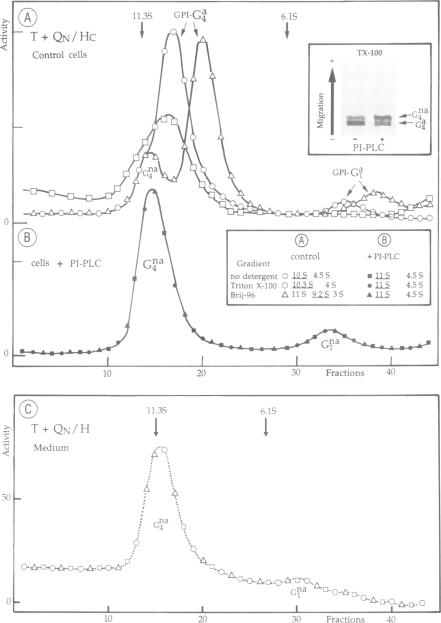
Asymmetric forms of Torpedo acetylcholinesterase (AChE) are produced in COS cells by the simultaneous expression of collagenic subunits (Q) and catalytic T subunits (AChET). Truncated AChET delta subunits, from which most of the C-terminal peptide (TC) had been deleted by mutagenesis, did not associate with Q subunits. The TC peptide is therefore necessary for the association of the AChET and Q subunits. In order to determine the orientation of the Q subunit in the collagen-tailed forms, we have developed an antiserum against its non-collagenic C-terminal domain, expressed as a fusion protein in Escherichia coli. This antiserum, which recognized the Q subunit in Western blots, was found to react with intact asymmetric forms, but not with collagenase-treated forms, from which the distal part of the tail had been cleaved, suggesting that the N-terminal non-collogenic domain (QN) is responsible for the interaction with the AChET subunits. This was confirmed by creating a chimeric subunit (QN/HC), in which QN was linked to the C-terminal peptide of the H subunit of Torpedo AChE, which contains the glycophosphatidylinositol (GPI) cleavage/attachment signal: co-expression of AChET and QN/NC produced GPI-anchored tetramers, which were sensitive to PI-PLC and largely exposed to the external surface of the cells. We thus demonstrate that: (i) the HC peptide is sufficient to determine the addition of a glycolipid anchor and (ii) the QN domain is sufficient to bind a catalytic AChET tetramer by interacting with the TC peptide.
通过同时表达胶原亚基(Q)和催化性T亚基(AChET),在COS细胞中产生了不对称形式的电鳐乙酰胆碱酯酶(AChE)。经诱变删除了大部分C末端肽(TC)的截短型AChET δ亚基,不与Q亚基结合。因此,TC肽对于AChET和Q亚基的结合是必需的。为了确定胶原尾形式中Q亚基的方向,我们制备了一种针对其非胶原C末端结构域的抗血清,该结构域在大肠杆菌中作为融合蛋白表达。这种抗血清在蛋白质免疫印迹中能识别Q亚基,发现它与完整的不对称形式反应,但不与胶原酶处理后的形式反应,后者的尾部远端部分已被切割,这表明N末端非胶原结构域(QN)负责与AChET亚基相互作用。通过构建嵌合亚基(QN/HC)证实了这一点,其中QN与电鳐AChE的H亚基的C末端肽相连,该C末端肽包含糖基磷脂酰肌醇(GPI)切割/附着信号:AChET和QN/NC共表达产生了GPI锚定的四聚体,它们对磷脂酰肌醇特异性磷脂酶C(PI-PLC)敏感,并且大部分暴露于细胞外表面。因此,我们证明:(i)HC肽足以决定糖脂锚的添加;(ii)QN结构域足以通过与TC肽相互作用结合催化性AChET四聚体。