Neuberger Georg, Kunze Markus, Eisenhaber Frank, Berger Johannes, Hartig Andreas, Brocard Cecile
Research Institute of Molecular Pathology (IMP), Dr Bohr-Gasse 7, A-1030 Vienna, Austria.
Genome Biol. 2004;5(12):R97. doi: 10.1186/gb-2004-5-12-r97. Epub 2004 Nov 30.
Can sequence segments coding for subcellular targeting or for posttranslational modifications occur in proteins that are not substrates in either of these processes? Although considerable effort has been invested in achieving low false-positive prediction rates, even accurate sequence-analysis tools for the recognition of these motifs generate a small but noticeable number of protein hits that lack the appropriate biological context but cannot be rationalized as false positives.
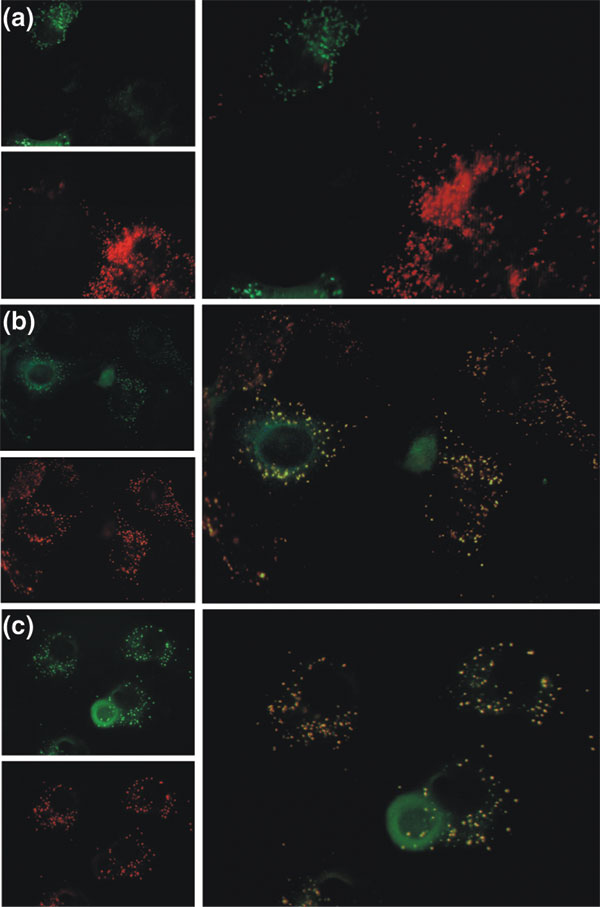
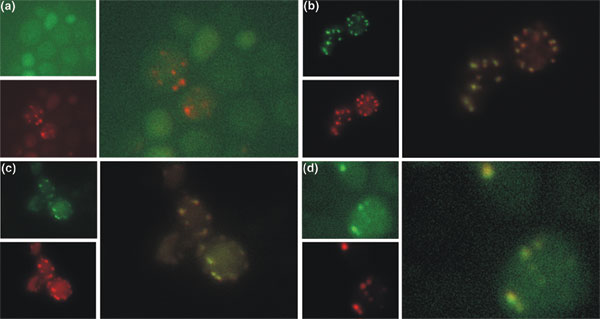
We show that the carboxyl termini of a set of definitely non-peroxisomal proteins with predicted peroxisomal targeting signals interact with the peroxisomal matrix protein receptor peroxin 5 (PEX5) in a yeast two-hybrid test. Moreover, we show that examples of these proteins - chicken lysozyme, human tyrosinase and the yeast mitochondrial ribosomal protein L2 (encoded by MRP7) - are imported into peroxisomes in vivo if their original sorting signals are disguised. We also show that even prokaryotic proteins can contain peroxisomal targeting sequences.
Thus, functional localization signals can evolve in unrelated protein sequences as a result of neutral mutations, and subcellular targeting is hierarchically organized, with signal accessibility playing a decisive role. The occurrence of silent functional motifs in unrelated proteins is important for the development of sequence-based function prediction tools and the interpretation of their results. Silent functional signals have the potential to acquire importance in future evolutionary scenarios and in pathological conditions.
编码亚细胞靶向或翻译后修饰的序列片段能否出现在并非这些过程中任何一个的底物的蛋白质中?尽管在实现低假阳性预测率方面投入了大量努力,但即使是用于识别这些基序的精确序列分析工具,也会产生少量但明显的蛋白质命中结果,这些结果缺乏适当的生物学背景,但又不能合理地解释为假阳性。
我们表明,一组具有预测的过氧化物酶体靶向信号的明确非过氧化物酶体蛋白质的羧基末端在酵母双杂交试验中与过氧化物酶体基质蛋白受体过氧化物酶5(PEX5)相互作用。此外,我们表明,如果这些蛋白质的原始分选信号被掩盖,它们的例子——鸡溶菌酶、人酪氨酸酶和酵母线粒体核糖体蛋白L2(由MRP7编码)——在体内会被导入过氧化物酶体。我们还表明,甚至原核生物蛋白质也可以包含过氧化物酶体靶向序列。
因此,由于中性突变,功能定位信号可以在不相关的蛋白质序列中进化,并且亚细胞靶向是分层组织的,信号可及性起决定性作用。不相关蛋白质中沉默功能基序的出现对于基于序列的功能预测工具的开发及其结果的解释很重要。沉默功能信号有可能在未来的进化场景和病理条件下变得重要。