Martin-DeLeon Patricia A, Zhang Hong, Morales Carlos R, Zhao Yutong, Rulon Michelle, Barnoski Barry L, Chen Hong, Galileo Deni S
Department of Biological Sciences, University of Delaware, Newark, DE 19716, USA.
Reprod Biol Endocrinol. 2005 Aug 10;3:32. doi: 10.1186/1477-7827-3-32.
While transmission ratio distortion, TRD, (a deviation from Mendelian ratio) is extensive in humans and well-documented in mice, the underlying mechanisms are unknown. Our earlier studies on carriers of spontaneous mutations of mouse Sperm Adhesion Molecule 1 (Spam1) suggested that TRD results from biochemically different sperm, due to a lack of transcript sharing through the intercellular cytoplasmic bridges of spermatids. These bridges usually allow transcript sharing among genetically different spermatids which develop into biochemically and functionally equivalent sperm.
The goals of the study were to provide support for the lack of sharing (LOS) hypothesis, using transgene and null carriers of Spam1, and to determine the mechanism of Spam1-associated TRD.
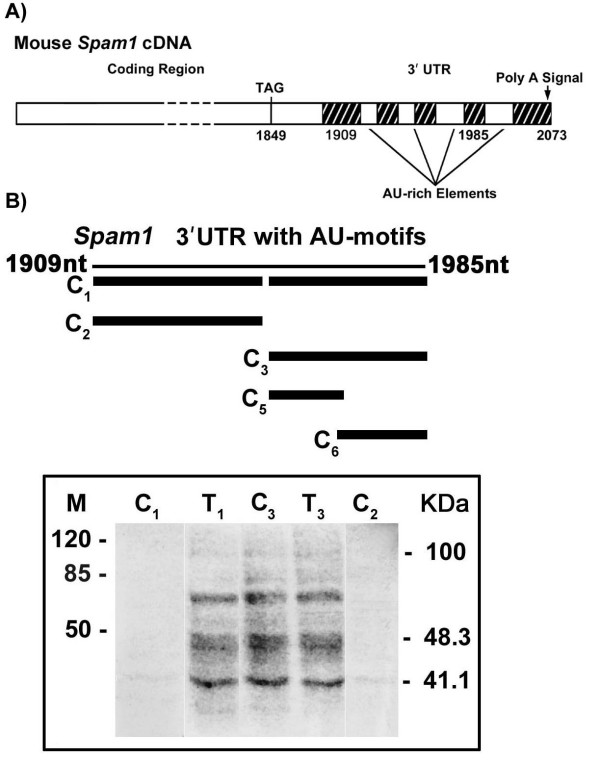
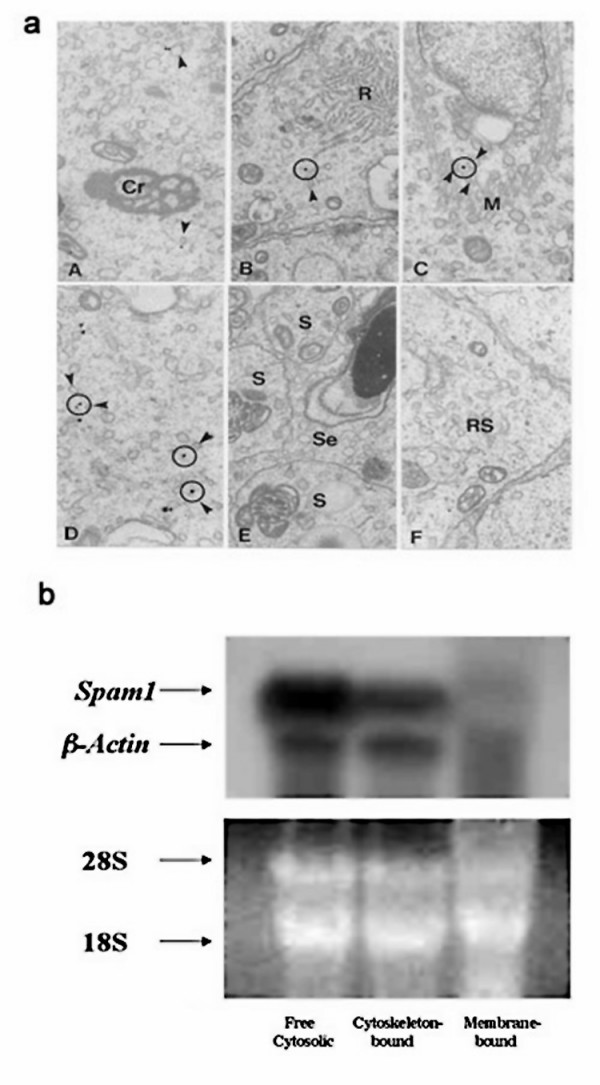
Carriers of Spam1-Hyal5 BAC transgenes were mated with wild-type female mice and the progeny analyzed for TRD by PCR genotyping. Sperm from transgene and Spam1 null carriers were analyzed using flow cytometry and immunocytochemistry to detect quantities of Spam1 and/or Hyal5. Transgene-bearing sperm with Spam1 overexpression were detected by fluorescence in situ hybridization. In wild-type animals, EM studies of in situ transcript hybridization of testis sections and Northern analysis of biochemically fractionated testicular RNA were performed to localize Spam1 transcript. Finally, AU-rich motifs identified in the 3' UTR of Spam1 RNA were assayed by UV cross-linking to determine their ability to interact with testicular RNA binding proteins.
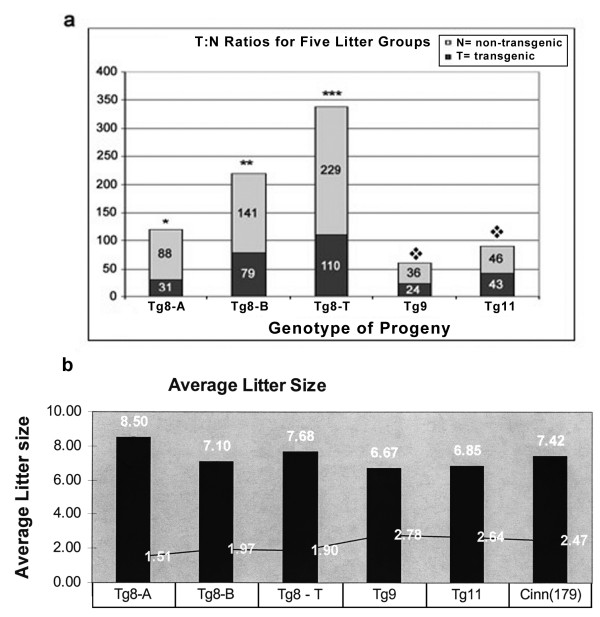
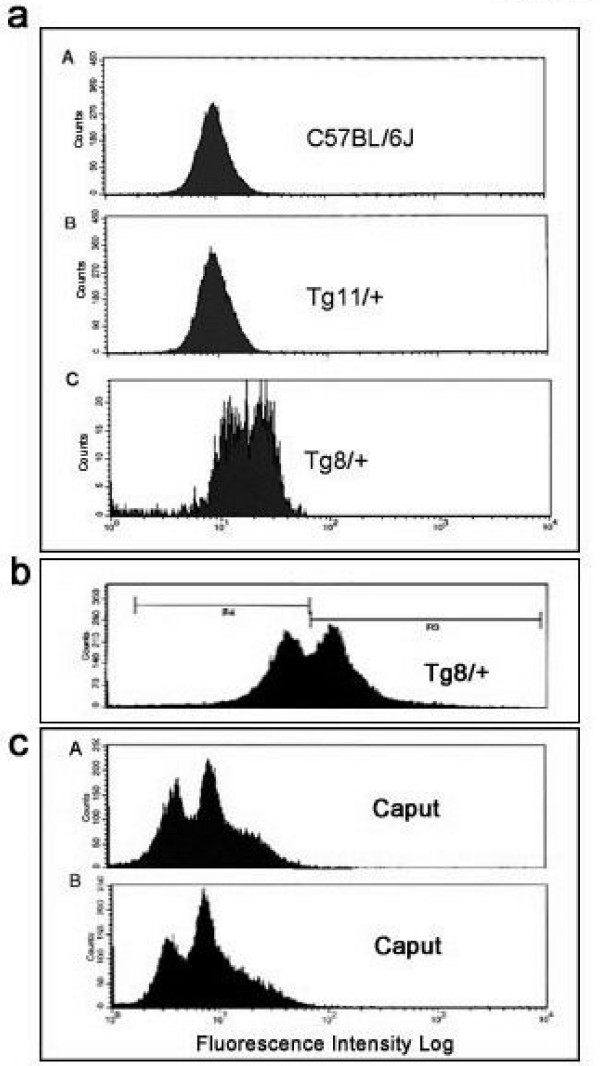
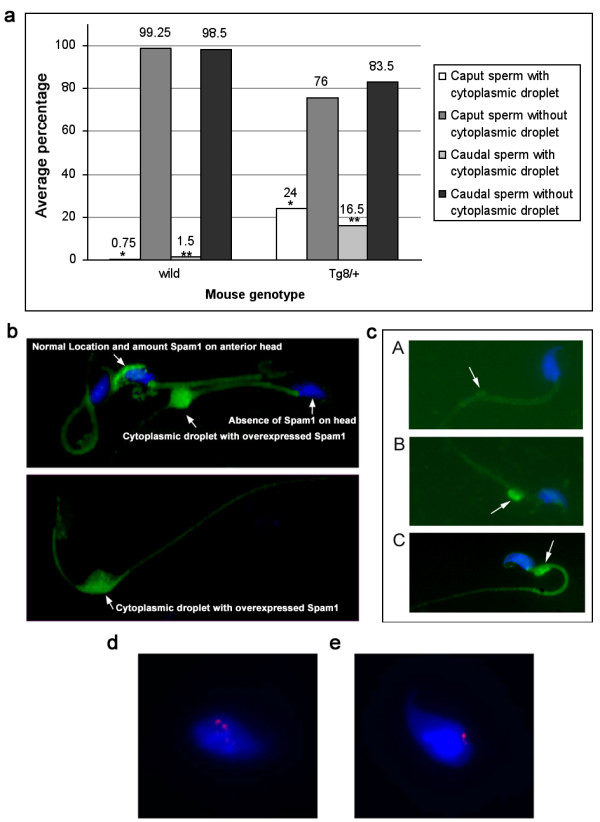
The Tg8 line of transgene carriers had a significant (P < 0.001) TRD, due to reduced fertilizing ability of transgene-bearing sperm. These sperm retained large cytoplasmic droplets engorged with overexpressed Spam1 or Hyal5 protein. Caudal sperm from transgene carriers and caput sperm of null carriers showed a bimodal distribution of Spam1, indicating that the sperm in a male were biochemically different with respect to Spam1 quantities. Spam1 RNA was absent from the bridges, associated exclusively with the ER, and was shown to be anchored to the cytoskeleton. This compartmentalization of the transcript, mediated by cytoskeletal binding, occurs via protein interactions with 3' UTR AU-rich sequences that are likely involved in its stabilization.
We provide strong support for the LOS hypothesis, and have elucidated the mechanism of Spam1-associated TRD.
虽然传递比失真(TRD,即偏离孟德尔比率)在人类中广泛存在且在小鼠中有充分记录,但其潜在机制尚不清楚。我们早期对小鼠精子粘附分子1(Spam1)自发突变携带者的研究表明,TRD是由于生化性质不同的精子所致,这是由于精子细胞间的胞质桥缺乏转录本共享。这些桥通常允许基因不同的精子细胞之间共享转录本,这些精子细胞发育成生化和功能上等效的精子。
本研究的目的是利用Spam1的转基因和基因敲除携带者为缺乏共享(LOS)假说提供支持,并确定与Spam1相关的TRD的机制。
将Spam1-Hyal5 BAC转基因携带者与野生型雌性小鼠交配,并通过PCR基因分型分析后代的TRD。使用流式细胞术和免疫细胞化学分析转基因和Spam1基因敲除携带者的精子,以检测Spam1和/或Hyal5的数量。通过荧光原位杂交检测过表达Spam1的转基因精子。在野生型动物中,对睾丸切片进行原位转录杂交的电镜研究,并对生化分级的睾丸RNA进行Northern分析,以定位Spam1转录本。最后,通过紫外线交联分析在Spam1 RNA的3'UTR中鉴定的富含AU的基序,以确定它们与睾丸RNA结合蛋白相互作用的能力。
转基因携带者的Tg8系存在显著(P < 0.001)的TRD,这是由于携带转基因的精子受精能力降低。这些精子保留了充满过表达的Spam1或Hyal5蛋白的大细胞质滴。转基因携带者的尾部精子和基因敲除携带者的头部精子显示出Spam1的双峰分布,表明雄性体内的精子在Spam1数量方面存在生化差异。胞质桥中不存在Spam1 RNA,其仅与内质网相关,并显示锚定在细胞骨架上。这种由细胞骨架结合介导的转录本分隔通过与3'UTR富含AU的序列的蛋白质相互作用发生,这些序列可能参与其稳定。
我们为LOS假说提供了有力支持,并阐明了与Spam1相关的TRD的机制。