Kobza Keyna, Camporeale Gabriela, Rueckert Brian, Kueh Alice, Griffin Jacob B, Sarath Gautam, Zempleni Janos
Department of Nutrition and Health Sciences, University of Nebraska-Lincoln, Lincoln, NE 68583-0806, USA.
FEBS J. 2005 Aug;272(16):4249-59. doi: 10.1111/j.1742-4658.2005.04839.x.
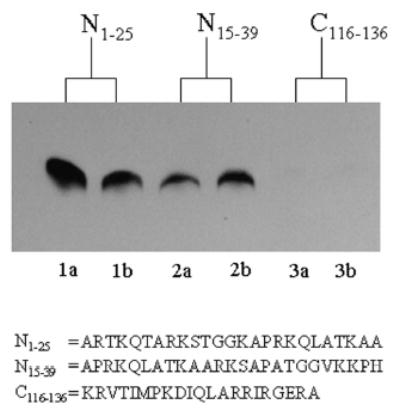
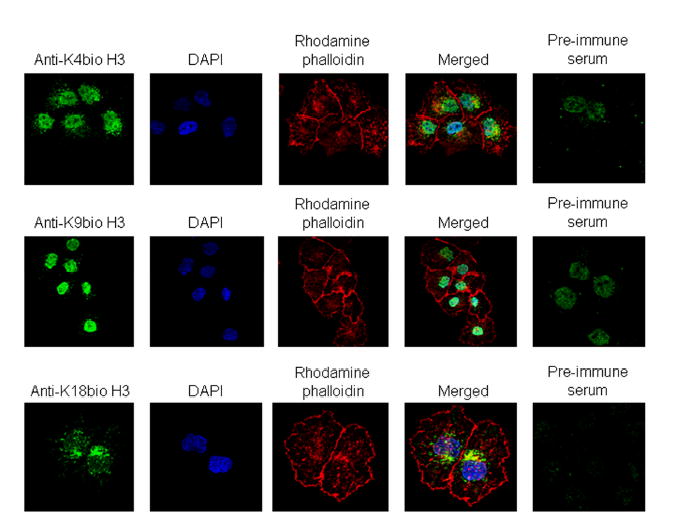
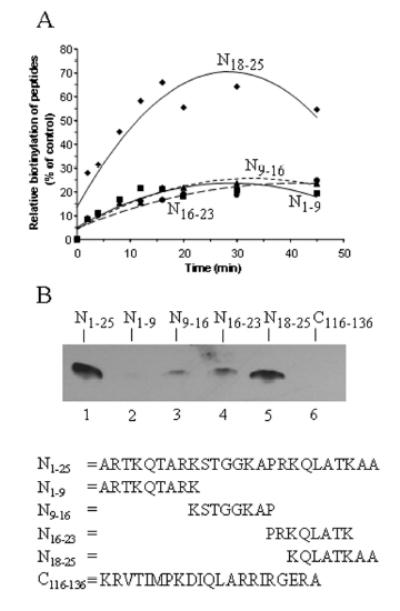
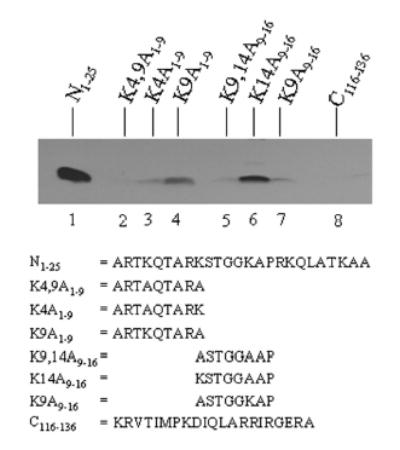
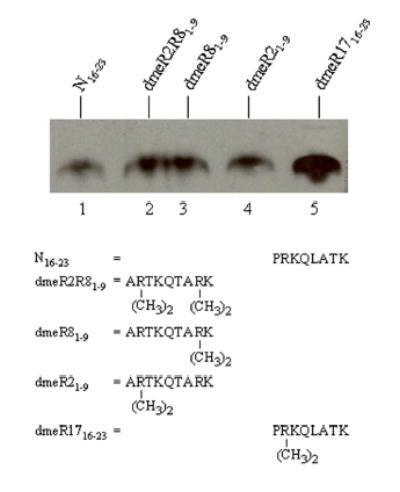
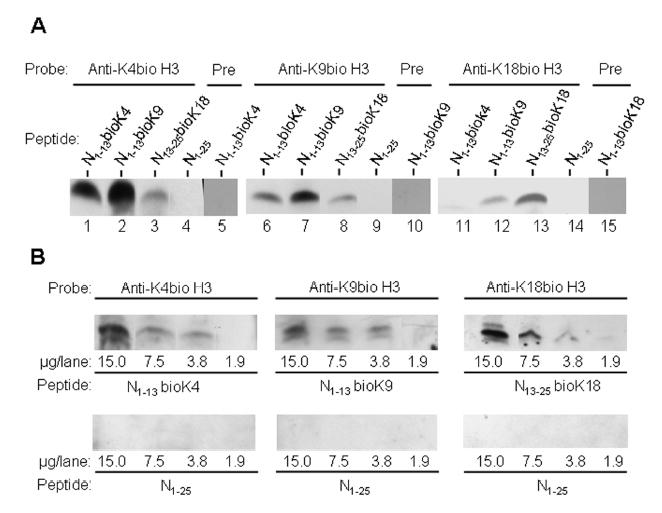
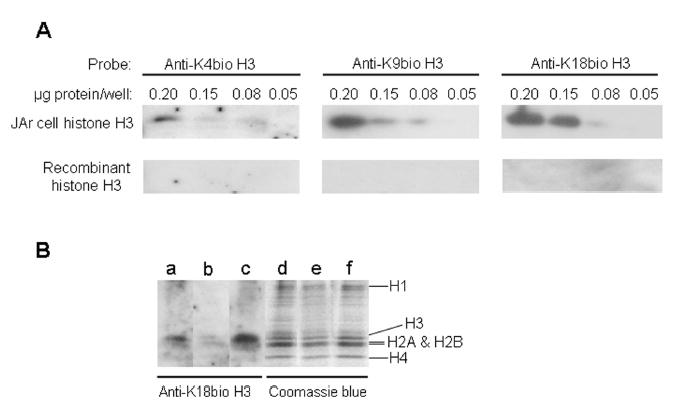
Histones are modified post-translationally, e.g. by methylation of lysine and arginine residues, and by phosphorylation of serine residues. These modifications regulate processes such as gene expression, DNA repair, and mitosis and meiosis. Recently, evidence has been provided that histones are also modified by covalent binding of the vitamin biotin. The aims of this study were to identify biotinylation sites in histone H3, and to investigate the crosstalk among histone biotinylation, methylation and phosphorylation. Synthetic peptides based on the sequence of human histone H3 were used as substrates for enzymatic biotinylation by biotinidase; biotin in peptides was probed using streptavidin peroxidase. These studies provided evidence that K4, K9 and K18 in histone H3 are good targets for biotinylation; K14 and K23 are relatively poor targets. Antibodies were generated to histone H3, biotinylated either at K4, K9 or K18. These antibodies localized to nuclei in human placental cells in immunocytochemistry and immunoblotting experiments, suggesting that lysines in histone H3 are biotinylated in vivo. Dimethylation of R2, R8 and R17 increased biotinylation of K4, K9 and K18, respectively, by biotinidase; phosphorylation of S10 abolished biotinylation of K9. These observations are consistent with crosstalk between biotinylation of histones and other known modifications of histones. We speculate that this crosstalk provides a link to known roles for biotin in gene expression and cell proliferation.
组蛋白在翻译后会发生修饰,例如赖氨酸和精氨酸残基的甲基化,以及丝氨酸残基的磷酸化。这些修饰调控着基因表达、DNA修复、有丝分裂和减数分裂等过程。最近,有证据表明组蛋白还会通过维生素生物素的共价结合进行修饰。本研究的目的是鉴定组蛋白H3中的生物素化位点,并研究组蛋白生物素化、甲基化和磷酸化之间的相互作用。基于人类组蛋白H3序列的合成肽被用作生物素酶进行酶促生物素化的底物;使用链霉抗生物素蛋白过氧化物酶检测肽中的生物素。这些研究提供了证据,表明组蛋白H3中的K4、K9和K18是生物素化的良好靶点;K14和K23是相对较差的靶点。制备了针对在K4、K9或K18处生物素化的组蛋白H3的抗体。在免疫细胞化学和免疫印迹实验中,这些抗体定位于人胎盘细胞的细胞核,表明组蛋白H3中的赖氨酸在体内被生物素化。R2、R8和R17的二甲基化分别增加了生物素酶对K4、K9和K18的生物素化;S10的磷酸化消除了K9的生物素化。这些观察结果与组蛋白生物素化和组蛋白其他已知修饰之间的相互作用一致。我们推测这种相互作用为生物素在基因表达和细胞增殖中的已知作用提供了联系。