Campbell Kirsteen J, O'Shea John M, Perkins Neil D
School of Life Sciences, Division of Gene Regulation and Expression, MSI/WTB, Complex, Dow Street, University of Dundee, Dundee, DD1 5EH, Scotland, UK.
BMC Cancer. 2006 Apr 21;6:101. doi: 10.1186/1471-2407-6-101.
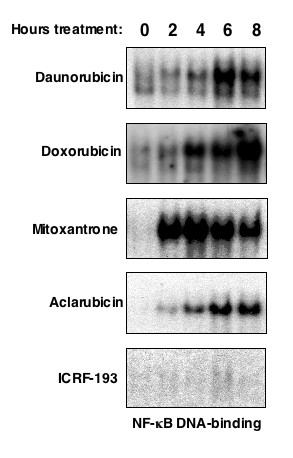
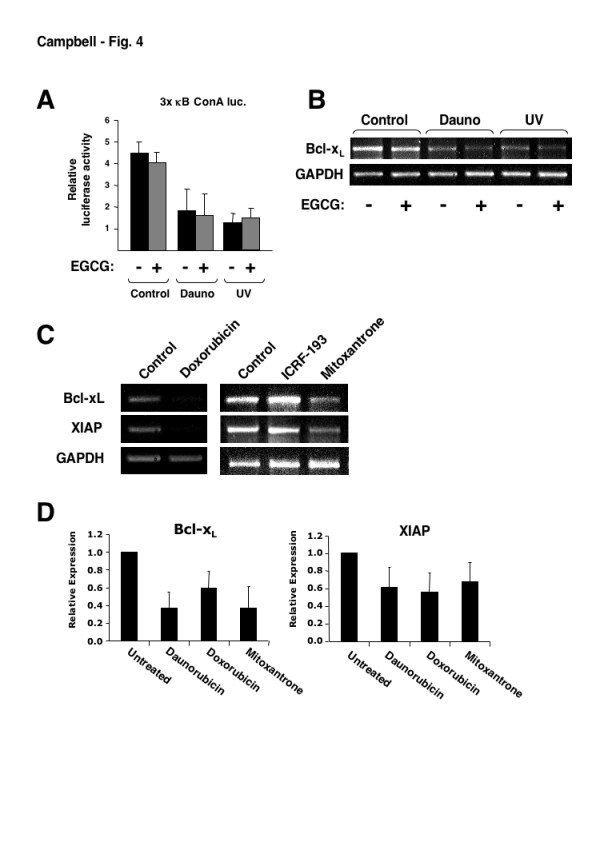
While many common chemotherapeutic drugs and other inducers of DNA-damage result in both NF-kappaB nuclear translocation and DNA-binding, we have previously observed that, depending on the precise stimulus, there is great diversity of the function of NF-kappaB. In particular, we found that treatment of U-2 OS osteosarcoma cells with the anthracycine daunorubicin or with ultraviolet (UV-C) light resulted in a form of NF-kappaB that repressed rather than induced NF-kappaB reporter plasmids and the expression of specific anti-apoptotic genes. Anthracyclines such as daunorubicin can induce DNA-damage though inhibiting topoisomerase II, intercalating with DNA and undergoing redox cycling to produce oxygen free radicals. In this study we have investigated other anthracyclines, doxorubicin and aclarubicin, as well as the anthracenedione mitoxantrone together with the topoisomerase II inhibitor ICRF-193, which all possess differing characteristics, to determine which of these features is specifically required to induce both NF-kappaB DNA-binding and transcriptional repression in U-2 OS cells.
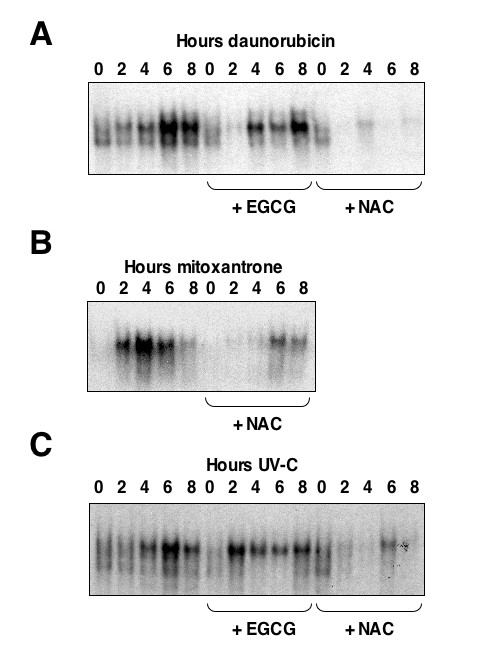
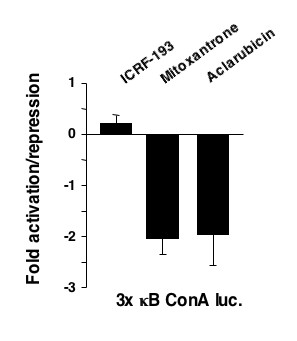
The use of mitoxantrone, which does not undergo redox cycling, and the reducing agent epigallocatechingallate (EGCG) demonstrated that oxygen free radical production is not required for induction of NF-kappaB DNA-binding and transcriptional repression by these agents and UV-C. In addition, the use of aclarubicin, which does not directly inhibit topoisomerase II and ICRF-193, which inhibits topoisomerase II but does not intercalate into DNA, demonstrated that topoisomerase II inhibition is not sufficient to induce the repressor form of NF-kappaB.
Induction of NF-kappaB DNA-binding and transcriptional repression by topoisomerase II inhibitors was found to correlate with an ability to intercalate into DNA. Although data from our and other laboratories indicates that topoisomerase II inhibition and oxygen free radicals do regulate NF-kappaB, they are not required for the particular ability of NF-kappaB to repress rather than activate transcription. Together with our previous data, these results demonstrate that the nature of the NF-kappaB response is context dependent. In a clinical setting such effects could profoundly influence the response to chemotherapy and suggest that new methods of analyzing NF-kappaB function could have both diagnostic and prognostic value.
虽然许多常见的化疗药物和其他DNA损伤诱导剂都会导致核因子κB(NF-κB)的核转位和DNA结合,但我们之前观察到,根据精确的刺激因素,NF-κB的功能存在很大差异。特别是,我们发现用蒽环类药物柔红霉素或紫外线(UV-C)处理U-2 OS骨肉瘤细胞会导致一种NF-κB形式,这种形式会抑制而非诱导NF-κB报告质粒以及特定抗凋亡基因的表达。像柔红霉素这样的蒽环类药物可通过抑制拓扑异构酶II、嵌入DNA以及进行氧化还原循环以产生氧自由基来诱导DNA损伤。在本研究中,我们研究了其他蒽环类药物阿霉素和阿柔比星,以及蒽二酮米托蒽醌,还有拓扑异构酶II抑制剂ICRF-193,它们都具有不同的特性,以确定在U-2 OS细胞中诱导NF-κB DNA结合和转录抑制具体需要这些特性中的哪一种。
使用不进行氧化还原循环的米托蒽醌以及还原剂表没食子儿茶素没食子酸酯(EGCG)表明,这些药物和UV-C诱导NF-κB DNA结合和转录抑制并不需要产生氧自由基。此外,使用不直接抑制拓扑异构酶II的阿柔比星和抑制拓扑异构酶II但不嵌入DNA的ICRF-193表明,抑制拓扑异构酶II不足以诱导NF-κB的抑制形式。
发现拓扑异构酶II抑制剂诱导NF-κB DNA结合和转录抑制与嵌入DNA的能力相关。尽管我们实验室和其他实验室的数据表明拓扑异构酶II抑制和氧自由基确实会调节NF-κB,但它们并非NF-κB抑制而非激活转录这一特定能力所必需。与我们之前的数据一起,这些结果表明NF-κB反应的性质取决于具体情况。在临床环境中,此类效应可能会深刻影响对化疗的反应,并表明分析NF-κB功能的新方法可能具有诊断和预后价值。