Division of Plant Breeding, Pioneer HiBred International Inc., Johnston, Iowa 50131.
Plant Physiol. 1990 Mar;92(3):755-60. doi: 10.1104/pp.92.3.755.
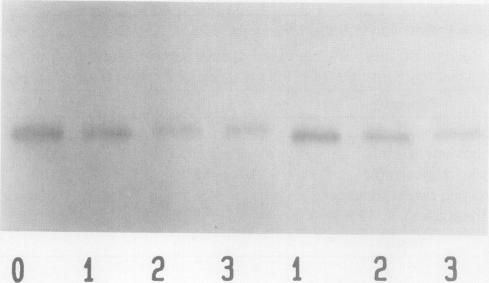
Respiration rates of Zea mays L. seedling tissues grown at 30 and 14 degrees C were measured at 25 degrees C at different stages of seedling growth. Accumulation of heat units was used to define the developmental stages to compare respiration between the two temperatures. At both temperatures, respiration rates of most tissues were highest at the youngest stages, then declined with age. Respiration rates of mesocotyl tissue were the most responsive to temperature, being nearly twofold higher when grown at 14 compared to 30 degrees C. Alternative pathway respiration increased concomitantly with respiration and was higher in mesocotyls grown in the cold. When seedlings were started at 30 then transferred to 14 degrees C, the increase in alternative pathway respiration due to cold was not observed unless the seedlings were transferred before 2 days of growth. Seedlings transferred to 14 degrees C after growth at 30 degrees C for 2 days had the same alternative oxidase capacity as seedlings grown at 30 degrees C. Seedlings grown at 14 degrees C for 10 to 12 days, then transferred to 30 degrees C, lost alternative pathway respiratory capacity over a period of 2 to 3 days. Western blots of mitochondrial proteins indicated that this loss of capacity was due to a loss of the alternative oxidase protein. Some in vitro characteristics of mitochondria were determined. The temperature optimum for measurement of alternative oxidase capacity was 15 to 20 degrees C. At 41 degrees C, very little alternative oxidase was measured, i.e., the mitochondrial oxygen uptake was almost completely sensitive to cyanide. This inactivation at 41 degrees C was reversible. After incubation at 41 degrees C, the alternative oxidase capacity measured at 25 degrees C was the similar to when it was measured at that temperature directly. Isolated mitochondria lost alternative oxidase capacity at the same rate when incubated at 41 degrees C as they did when incubated at 25 degrees C. Increasing the supply of electrons to isolated mitochondria increased the degree of engagement of the alternative pathway, whereas lower temperature decreased the degree of engagement. Lower temperatures did not increase the degree of engagement of the pathway in intact tissues. We interpret these observations to indicate that the greater capacity of alternative oxidase in cold-grown seedlings is a consequence of development at these low temperatures which results in elevated respiration rates. Low temperature itself does not cause greater capacity or engagement of the alternative oxidase in mitochondria that have developed under warm temperatures. Our hypothesis would be that the low growth temperatures require the seedlings to have a higher respiration rate for some reason, e.g., to prevent the accumulation of a toxic metabolite, and that the alternative pathway functions in that respiration.
在 30°C 和 14°C 下生长的玉米幼苗组织在 25°C 下的不同生长阶段的呼吸速率进行了测量。用热单位的积累来定义发育阶段,以比较两种温度下的呼吸作用。在这两种温度下,大多数组织的呼吸速率在最年轻的阶段最高,然后随着年龄的增长而下降。中胚轴组织的呼吸速率对温度的反应最为敏感,在 14°C 下生长时比在 30°C 下生长时高出近两倍。替代途径的呼吸作用与呼吸作用同时增加,在寒冷条件下生长的中胚轴中更高。当幼苗在 30°C 下开始生长,然后转移到 14°C 时,如果幼苗在生长 2 天之前没有转移,就不会观察到由于低温引起的替代途径呼吸作用的增加。在 30°C 下生长 2 天后转移到 14°C 的幼苗,其替代氧化酶的能力与在 30°C 下生长的幼苗相同。在 14°C 下生长 10 至 12 天,然后转移到 30°C 的幼苗在 2 至 3 天内失去了替代途径的呼吸能力。线粒体蛋白的 Western 印迹表明,这种能力的丧失是由于替代氧化酶蛋白的丧失。还确定了一些线粒体的体外特性。测量替代氧化酶能力的最佳温度为 15°C 至 20°C。在 41°C 时,测量到的替代氧化酶很少,即线粒体耗氧量几乎完全对氰化物敏感。这种在 41°C 下的失活是可逆的。在 41°C 下孵育后,在 25°C 下测量的替代氧化酶能力与直接在该温度下测量的能力相似。在 41°C 下孵育时,与在 25°C 下孵育时相比,分离的线粒体以相同的速度失去替代氧化酶能力。向分离的线粒体中增加电子供应会增加替代途径的参与程度,而较低的温度会降低参与程度。较低的温度不会增加完整组织中途径的参与程度。我们的观察结果表明,在低温下生长的幼苗中替代氧化酶的更大能力是在这些低温下发育的结果,导致呼吸速率升高。低温本身不会导致在温暖温度下发育的线粒体中替代氧化酶的更大能力或参与度。我们的假设是,低温生长需要幼苗以某种方式具有更高的呼吸速率,例如,防止有毒代谢物的积累,并且替代途径在该呼吸中起作用。