Kaplan J M, Varmus H E, Bishop J M
G.W. Hooper Research Foundation, University of California Medical Center, San Francisco 94143.
Mol Cell Biol. 1990 Mar;10(3):1000-9. doi: 10.1128/mcb.10.3.1000-1009.1990.
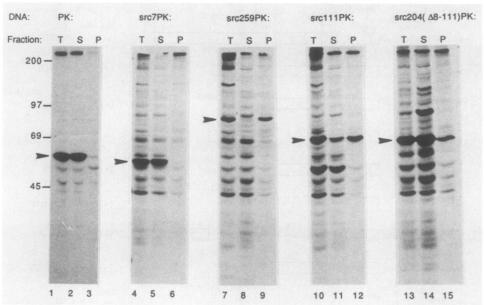
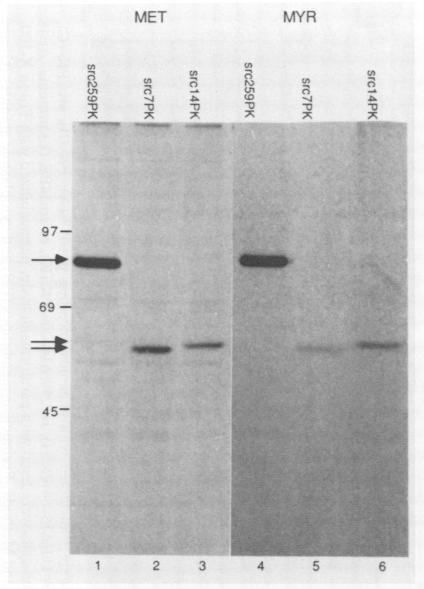
The proteins encoded by the oncogene v-src and its cellular counterpart c-src (designated generically here as pp60src) are tightly associated with both plasma membranes and intracellular membranes. This association is due in part to the amino-terminal myristylation of pp60src, but several lines of evidence suggest that amino-terminal portions of the protein itself are also involved. We now report that pp60src contains at least three domains which, in conjunction with myristylation, are capable of mediating attachment to membranes and determining subcellular localization. We identified these domains by fusing various portions of pp60src to pyruvate kinase, which is normally a cytoplasmic protein. Amino acids 1 to 14 of pp60src are sufficient to mediate both myristylation and the attachment of pyruvate kinase to cytoplasmic granules. In contrast, amino acids 38 to 111 mediate association with the plasma membrane and perinuclear membranes, whereas amino acids 204 to 259 mediate association primarily with perinuclear membranes. We conclude that pp60src contains independent domains that target the protein to distinctive subcellular locations and thus may facilitate diverse biological functions of the protein.
癌基因v-src及其细胞对应物c-src(此处统称为pp60src)编码的蛋白质与质膜和细胞内膜紧密相关。这种关联部分归因于pp60src的氨基末端肉豆蔻酰化,但有几条证据表明该蛋白质本身的氨基末端部分也参与其中。我们现在报告,pp60src包含至少三个结构域,这些结构域与肉豆蔻酰化一起,能够介导与膜的附着并确定亚细胞定位。我们通过将pp60src的各个部分与丙酮酸激酶融合来鉴定这些结构域,丙酮酸激酶通常是一种细胞质蛋白。pp60src的第1至14位氨基酸足以介导肉豆蔻酰化以及丙酮酸激酶与细胞质颗粒的附着。相比之下,第38至111位氨基酸介导与质膜和核周膜的关联,而第204至259位氨基酸主要介导与核周膜的关联。我们得出结论,pp60src包含独立的结构域,这些结构域将该蛋白质靶向不同的亚细胞位置,因此可能促进该蛋白质的多种生物学功能。