He Quanhua, Alexeev Dmitriy, Estevez Maureen E, McCabe Sarah L, Calvert Peter D, Ong David E, Cornwall M Carter, Zimmerman Anita L, Makino Clint L
Department of Ophthalmology, Massachusetts Eye and Ear Infirmary and Harvard Medical School, Boston, MA 02114, USA.
J Gen Physiol. 2006 Oct;128(4):473-85. doi: 10.1085/jgp.200609619.
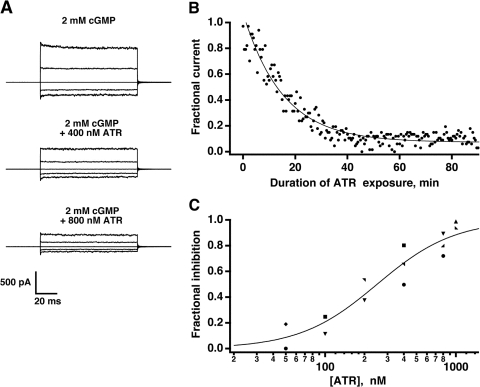
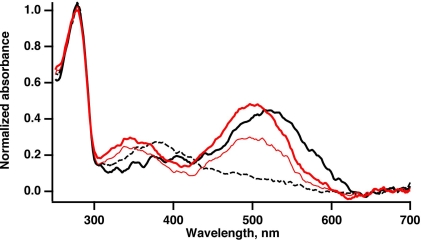
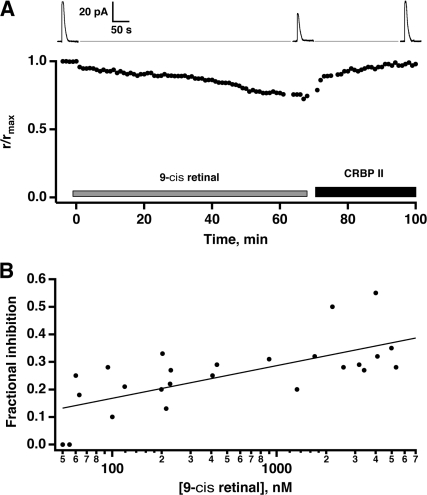
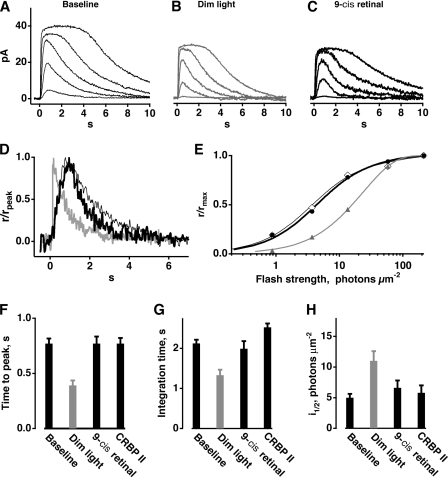
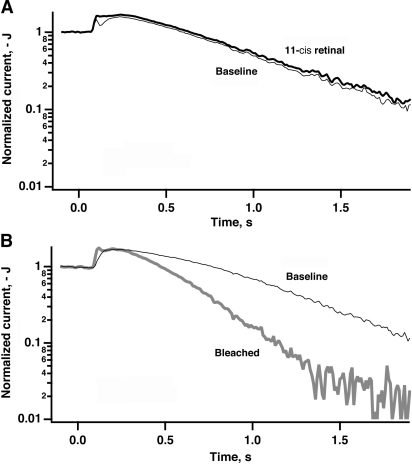
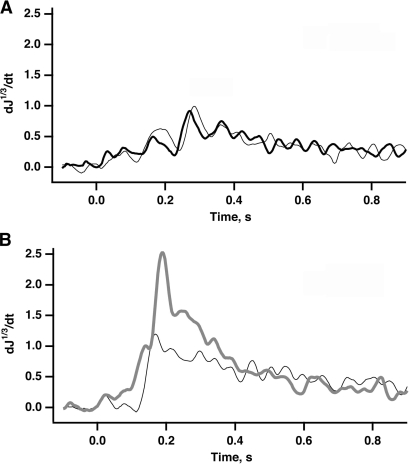
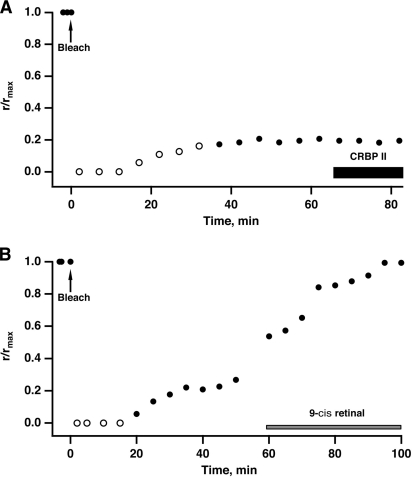
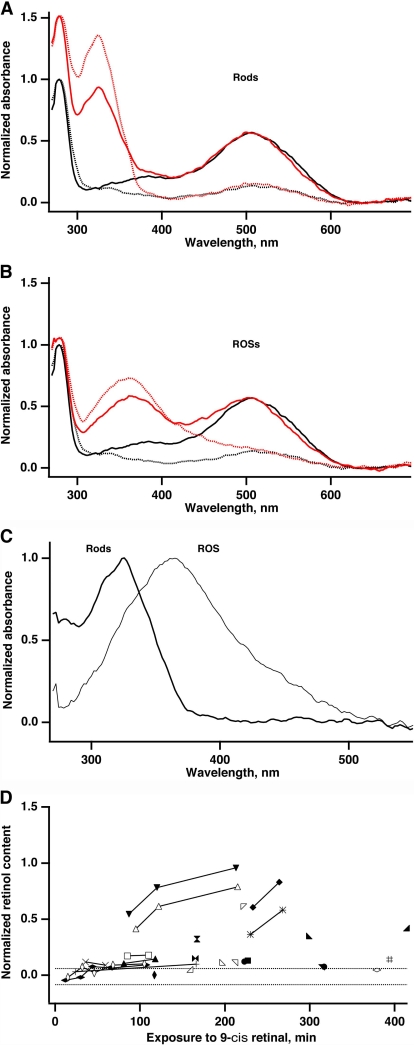
In vertebrate rods, photoisomerization of the 11-cis retinal chromophore of rhodopsin to the all-trans conformation initiates a biochemical cascade that closes cGMP-gated channels and hyperpolarizes the cell. All-trans retinal is reduced to retinol and then removed to the pigment epithelium. The pigment epithelium supplies fresh 11-cis retinal to regenerate rhodopsin. The recent discovery that tens of nanomolar retinal inhibits cloned cGMP-gated channels at low [cGMP] raised the question of whether retinoid traffic across the plasma membrane of the rod might participate in the signaling of light. Native channels in excised patches from rods were very sensitive to retinoid inhibition. Perfusion of intact rods with exogenous 9- or 11-cis retinal closed cGMP-gated channels but required higher than expected concentrations. Channels reopened after perfusing the rod with cellular retinoid binding protein II. PDE activity, flash response kinetics, and relative sensitivity were unchanged, ruling out pharmacological activation of the phototransduction cascade. Bleaching of rhodopsin to create all-trans retinal and retinol inside the rod did not produce any measurable channel inhibition. Exposure of a bleached rod to 9- or 11-cis retinal did not elicit channel inhibition during the period of rhodopsin regeneration. Microspectrophotometric measurements showed that exogenous 9- or 11-cis retinal rapidly cross the plasma membrane of bleached rods and regenerate their rhodopsin. Although dark-adapted rods could also take up large quantities of 9-cis retinal, which they converted to retinol, the time course was slow. Apparently cGMP-gated channels in intact rods are protected from the inhibitory effects of retinoids that cross the plasma membrane by a large-capacity buffer. Opsin, with its chromophore binding pocket occupied (rhodopsin) or vacant, may be an important component. Exceptionally high retinoid levels, e.g., associated with some retinal degenerations, could overcome the buffer, however, and impair sensitivity or delay the recovery after exposure to bright light.
在脊椎动物的视杆细胞中,视紫红质的11-顺式视黄醛发色团光异构化为全反式构象,引发了一系列生化反应,导致环鸟苷酸门控通道关闭,细胞超极化。全反式视黄醛被还原为视黄醇,然后被转运至色素上皮。色素上皮提供新鲜的11-顺式视黄醛以再生视紫红质。最近发现,在低[cGMP]浓度下,数十纳摩尔的视黄醛会抑制克隆的环鸟苷酸门控通道,这引发了一个问题,即视黄醛穿过视杆细胞质膜的运输是否可能参与光信号传导。从视杆细胞分离的膜片中的天然通道对视黄醛抑制非常敏感。用外源性9-或11-顺式视黄醛灌注完整的视杆细胞会关闭环鸟苷酸门控通道,但所需浓度高于预期。在用细胞视黄醛结合蛋白II灌注视杆细胞后,通道重新开放。磷酸二酯酶(PDE)活性、闪光反应动力学和相对敏感性均未改变,排除了光转导级联反应的药理学激活。视紫红质漂白在视杆细胞内产生全反式视黄醛和视黄醇,并未产生任何可测量的通道抑制。在视紫红质再生期间,将漂白的视杆细胞暴露于9-或11-顺式视黄醛不会引发通道抑制。显微分光光度测量表明,外源性9-或11-顺式视黄醛可迅速穿过漂白视杆细胞的质膜并再生其视紫红质。尽管暗适应的视杆细胞也能摄取大量的9-顺式视黄醛并将其转化为视黄醇,但其时间进程较慢。显然,完整视杆细胞中的环鸟苷酸门控通道受到一种大容量缓冲剂的保护,免受穿过质膜的视黄醛的抑制作用。视蛋白,无论其发色团结合口袋是被占据(视紫红质)还是空的,可能都是一个重要组成部分。然而,例如与某些视网膜变性相关的异常高视黄醛水平可能会克服这种缓冲作用,损害敏感性或延迟暴露于强光后的恢复。