Cohen Roy, Marom Merav, Atlas Daphne
Department of Biological Chemistry, The Alexander Silberman Institute of Life Sciences, The Hebrew University of Jerusalem, Jerusalem, Israel.
PLoS One. 2007 Dec 5;2(12):e1273. doi: 10.1371/journal.pone.0001273.
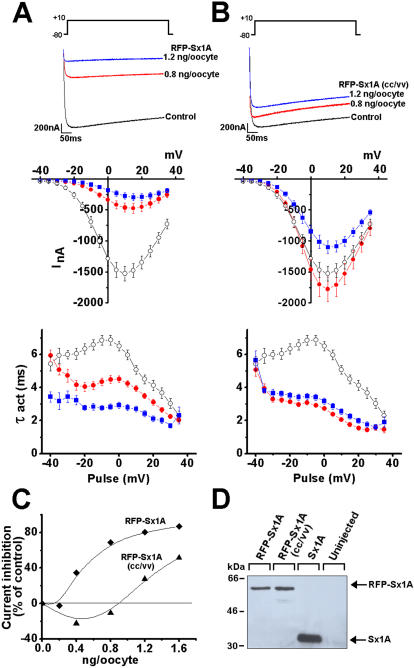
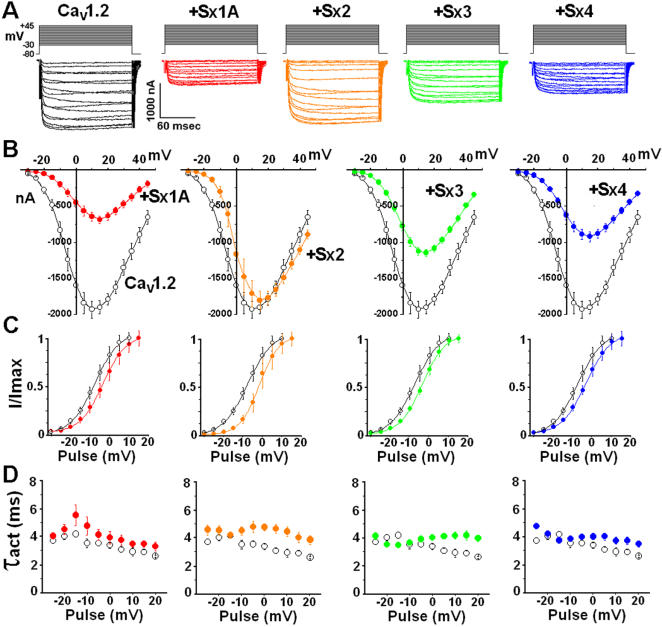
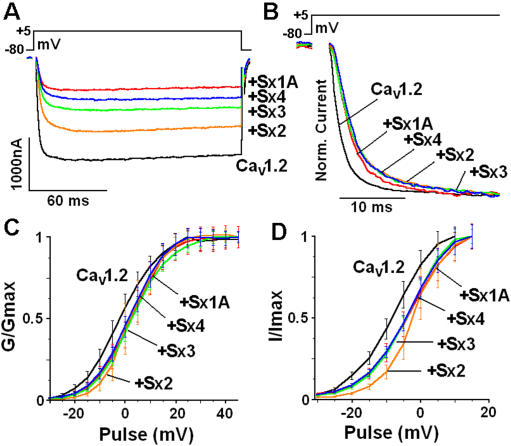
The interactions of the voltage-gated Ca(2+) channel (VGCC) with syntaxin 1A (Sx 1A), Synaptosome-associated protein of 25 kD (SNAP-25), and synaptotagmin, couple electrical excitation to evoked secretion. Two vicinal Cys residues, Cys 271 and Cys 272 in the Sx 1A transmembrane domain, are highly conserved and participate in modulating channel kinetics. Each of the Sx1A Cys mutants, differently modify the kinetics of Cav1.2, and neuronal Cav2.2 calcium channel.
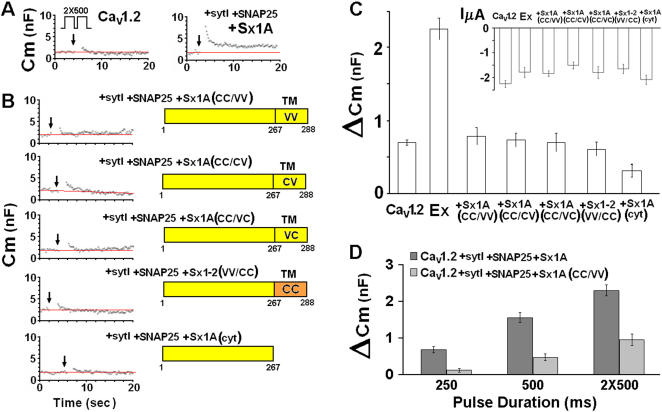
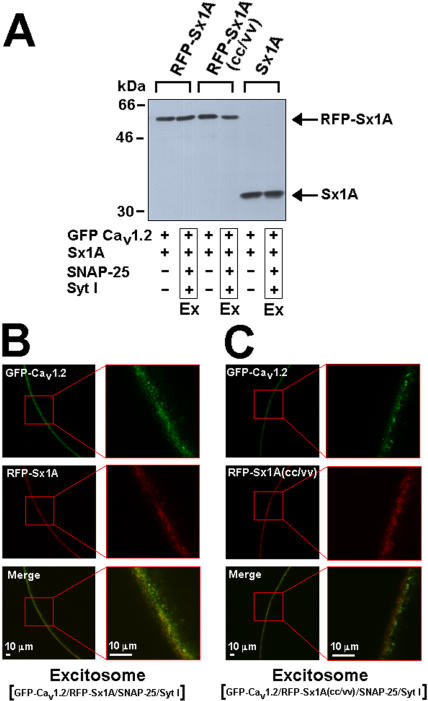
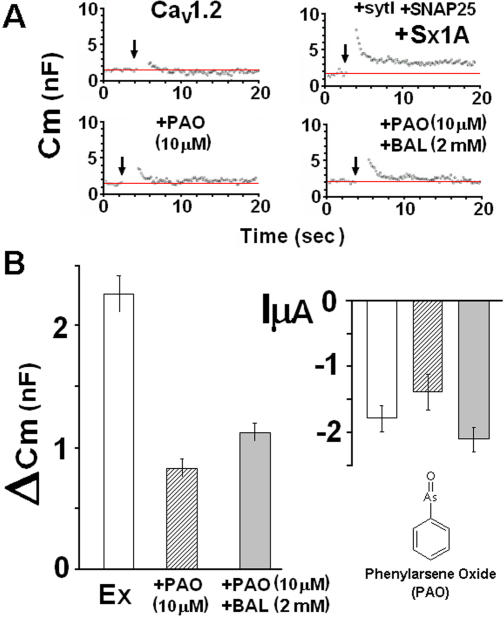
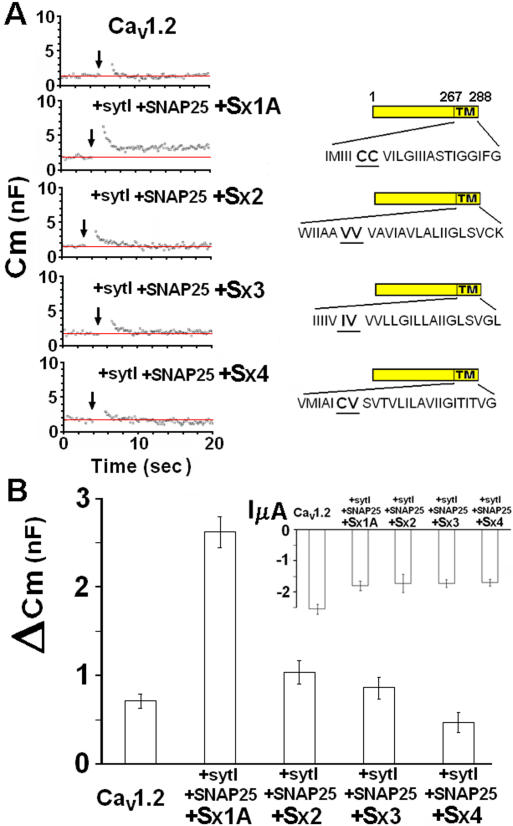
METHODOLOGY/PRINCIPLE FINDINGS: We examined the effects of various Sx1A Cys mutants and the syntaxin isoforms 2, 3, and 4 each of which lack vicinal Cys residues, on evoked secretion, monitoring capacitance transients in a functional release assay. Membrane capacitance in Xenopus oocytes co-expressing Cav1.2, Sx1A, SNAP-25 and synaptotagmin, which is Bot C- and Bot A-sensitive, was elicited by a double 500 ms depolarizing pulse to 0 mV. The evoked-release was obliterated when a single Cys Sx1A mutant or either one of the Sx isoforms were substituted for Sx 1A, demonstrating the essential role of vicinal Cys residues in the depolarization mediated process. Protein expression and confocal imaging established the level of the mutated proteins in the cell and their targeting to the plasma membrane.
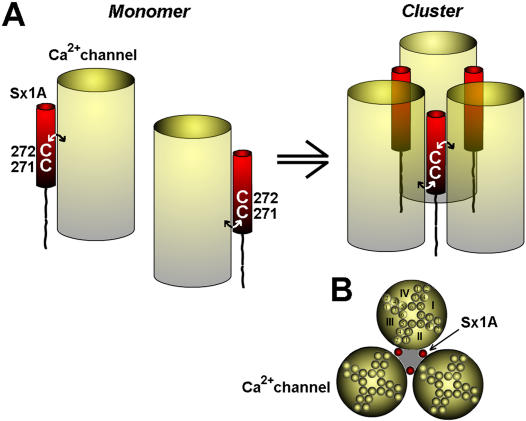
CONCLUSIONS/SIGNIFICANCE: We propose a model whereby the two adjacent transmembranal Cys residues of Sx 1A, lash two calcium channels. Consistent with the necessity of a minimal fusion complex termed the excitosome, each Sx1A is in a complex with SNAP-25, Syt1, and the Ca(2+) channel. A Hill coefficient >2 imply that at least three excitosome complexes are required for generating a secreting hetero-oligomer protein complex. This working model suggests that a fusion pore that opens during membrane depolarization could be lined by alternating transmembrane segments of Sx1A and VGCC. The functional coupling of distinct amino acids of Sx 1A with VGCC appears to be essential for depolarization-evoked secretion.
电压门控钙通道(VGCC)与 syntaxin 1A(Sx 1A)、25kD 的突触小体相关蛋白(SNAP - 25)以及突触结合蛋白相互作用,将电兴奋与诱发分泌偶联起来。Sx 1A 跨膜结构域中的两个相邻半胱氨酸残基,即 Cys 271 和 Cys 272,高度保守并参与调节通道动力学。每个 Sx1A 半胱氨酸突变体对 Cav1.2 和神经元 Cav2.2 钙通道的动力学有不同程度的改变。
方法/主要发现:我们在功能释放实验中监测电容瞬变,研究了各种 Sx1A 半胱氨酸突变体以及缺乏相邻半胱氨酸残基的 syntaxin 同工型 2、3 和 4 对诱发分泌的影响。通过对共表达 Cav1.2、Sx1A、SNAP - 25 和突触结合蛋白(对肉毒杆菌 C 和肉毒杆菌 A 敏感)的非洲爪蟾卵母细胞施加两个 500 毫秒的去极化脉冲至 0 mV 来引发膜电容变化。当单个半胱氨酸 Sx1A 突变体或任何一种 Sx 同工型替代 Sx 1A时,诱发释放被消除,这表明相邻半胱氨酸残基在去极化介导的过程中起着至关重要的作用。蛋白质表达和共聚焦成像确定了细胞中突变蛋白的水平及其靶向质膜的情况。
结论/意义:我们提出了一个模型,即 Sx 1A 的两个相邻跨膜半胱氨酸残基与两个钙通道相连。与称为兴奋体的最小融合复合物的必要性一致,每个 Sx1A 都与 SNAP - 25、Syt1 和 Ca(2+)通道形成复合物。希尔系数>2 意味着至少需要三个兴奋体复合物来生成一个分泌性异源寡聚体蛋白复合物。这个工作模型表明,在膜去极化过程中打开的融合孔可能由 Sx1A 和 VGCC 的交替跨膜片段构成。Sx 1A 的不同氨基酸与 VGCC 的功能偶联似乎对去极化诱发的分泌至关重要。