Siepka Sandra M, Yoo Seung-Hee, Park Junghea, Lee Choogon, Takahashi Joseph S
Howard Hughes Medical Institute, Northwestern University 2205 Tech Drive Evanston, IL 60208, USA.
Center for Functional Genomics, Northwestern University 2205 Tech Drive Evanston, IL 60208, USA.
Cold Spring Harb Symp Quant Biol. 2007;72:251-259. doi: 10.1101/sqb.2007.72.052.
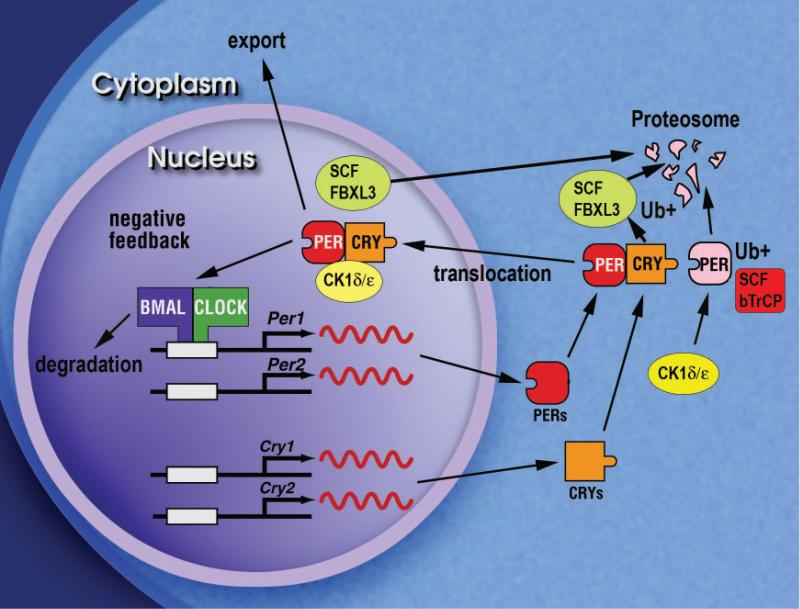
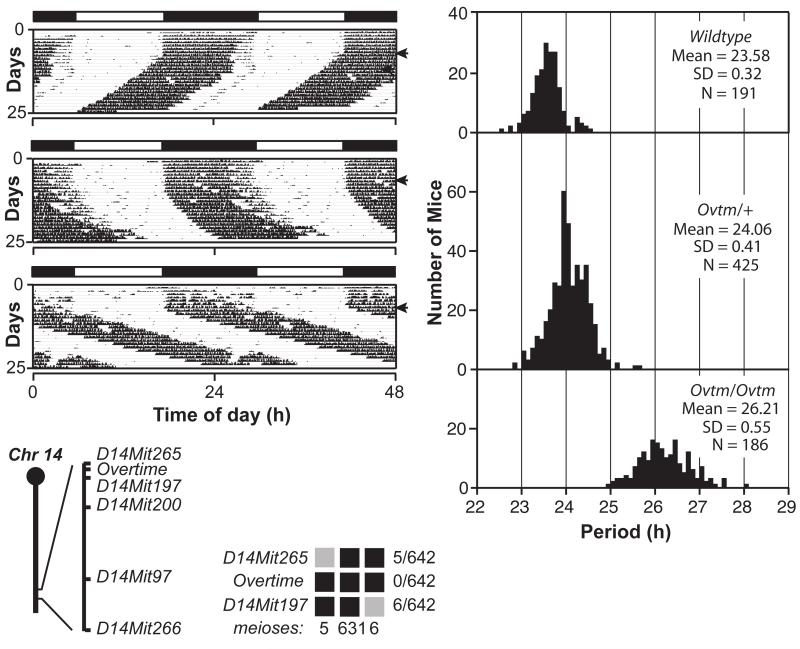
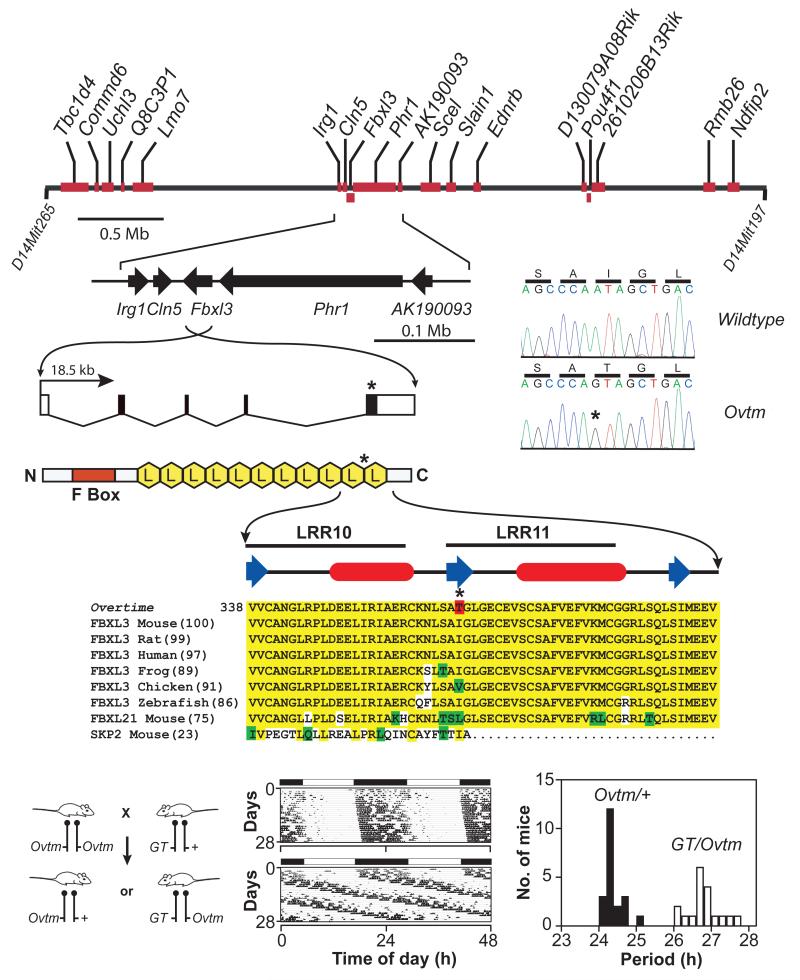
In animals, circadian behavior can be analyzed as an integrated system, beginning with genes and leading ultimately to behavioral outputs. In the last decade, the molecular mechanism of circadian clocks has been unraveled primarily by the use of phenotype-driven (forward) genetic analysis in a number of model systems. Circadian oscillations are generated by a set of genes forming a transcriptional autoregulatory feedback loop. In mammals, there is a "core" set of circadian genes that form the primary negative feedback loop of the clock mechanism (Clock/Npas2, Bmal1, Per1, Per2, Cry1, Cry2, and CK1epsilon). A further dozen candidate genes have been identified and have additional roles in the circadian gene network such as the feedback loop involving Rev-erbalpha. Despite this remarkable progress, it is clear that a significant number of genes that strongly influence and regulate circadian rhythms in mammals remain to be discovered and identified. As part of a large-scale N-ethyl-N-nitrosourea mutagenesis screen using a wide range of nervous system and behavioral phenotypes, we have identified a number of new circadian mutants in mice. Here, we describe a new short-period circadian mutant, part-time (prtm), which is caused by a loss-of-function mutation in the Cryptochrome1 (Cry1) gene. We also describe a long-period circadian mutant named Overtime (Ovtm). Positional cloning and genetic complementation reveal that Ovtm is encoded by the F-box protein FBXL3, a component of the SKP1-CUL1-F-box protein (SCF) E3 ubiquitin ligase complex. The Ovtm mutation causes an isoleucine to threonine (I364T) substitution leading to a loss of function in FBXL3 that interacts specifically with the CRYPTOCHROME (CRY) proteins. In Ovtm mice, expression of the PERIOD proteins PER1 and PER2 is reduced; however, the CRY proteins CRY1 and CRY2 are unchanged. The loss of FBXL3 function leads to a stabilization of the CRY proteins, which in turn leads to a global transcriptional repression of the Per and Cry genes. Thus, Fbxl3(Ovtm) defines a molecular link between CRY turnover and CLOCK/BMAL1-dependent circadian transcription to modulate circadian period.
在动物中,昼夜节律行为可作为一个整合系统进行分析,始于基因并最终导致行为输出。在过去十年中,昼夜节律时钟的分子机制主要通过在一些模型系统中使用表型驱动(正向)遗传分析得以阐明。昼夜节律振荡由一组形成转录自调节反馈环的基因产生。在哺乳动物中,有一组“核心”昼夜节律基因形成时钟机制的主要负反馈环(Clock/Npas2、Bmal1、Per1、Per2、Cry1、Cry2和CK1epsilon)。另外还鉴定出了十几个候选基因,它们在昼夜节律基因网络中具有其他作用,例如涉及Rev-erbalpha的反馈环。尽管取得了这一显著进展,但显然仍有大量强烈影响和调节哺乳动物昼夜节律的基因有待发现和鉴定。作为使用广泛的神经系统和行为表型进行的大规模N-乙基-N-亚硝基脲诱变筛选的一部分,我们在小鼠中鉴定出了一些新的昼夜节律突变体。在此,我们描述了一种新的短周期昼夜节律突变体——兼职(prtm),它是由隐花色素1(Cry1)基因的功能丧失突变引起的。我们还描述了一种名为加班(Ovtm)的长周期昼夜节律突变体。定位克隆和遗传互补表明,Ovtm由F-box蛋白FBXL3编码,FBXL3是SKP1-CUL1-F-box蛋白(SCF)E3泛素连接酶复合物的一个组成部分。Ovtm突变导致异亮氨酸到苏氨酸(I364T)的替换,导致与隐花色素(CRY)蛋白特异性相互作用的FBXL3功能丧失。在Ovtm小鼠中,周期蛋白PER1和PER2的表达降低;然而,CRY蛋白CRY1和CRY2没有变化。FBXL3功能的丧失导致CRY蛋白的稳定,这反过来又导致Per和Cry基因的全局转录抑制。因此,Fbxl3(Ovtm)定义了CRY周转与CLOCK/BMAL1依赖性昼夜节律转录之间的分子联系,以调节昼夜节律周期。