Merks Roeland M H, Perryn Erica D, Shirinifard Abbas, Glazier James A
VIB Department of Plant Systems Biology, Ghent, Belgium.
PLoS Comput Biol. 2008 Sep 19;4(9):e1000163. doi: 10.1371/journal.pcbi.1000163.
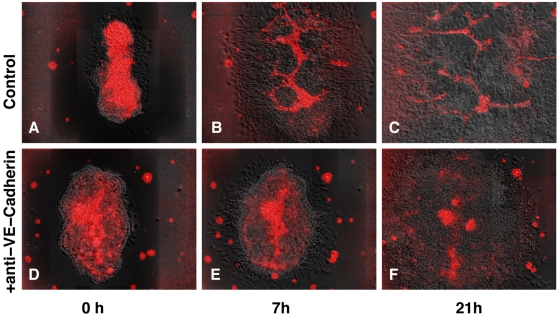
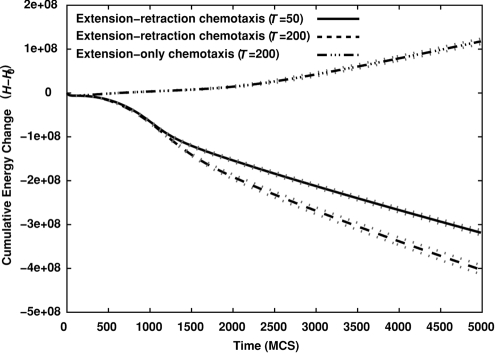
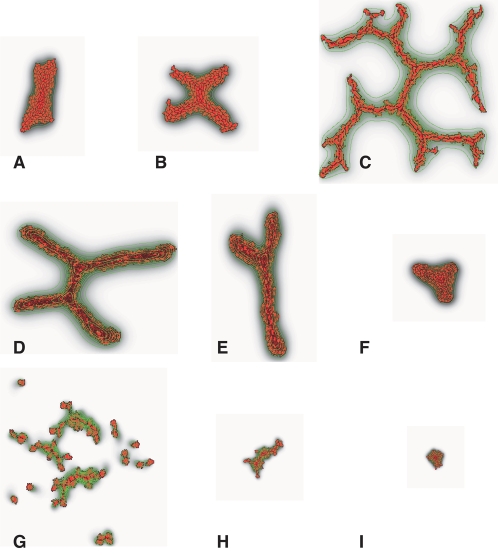
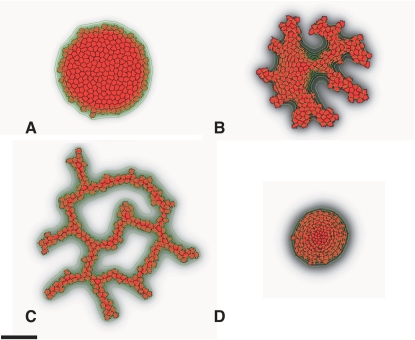
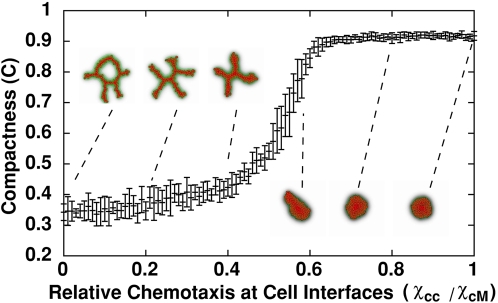
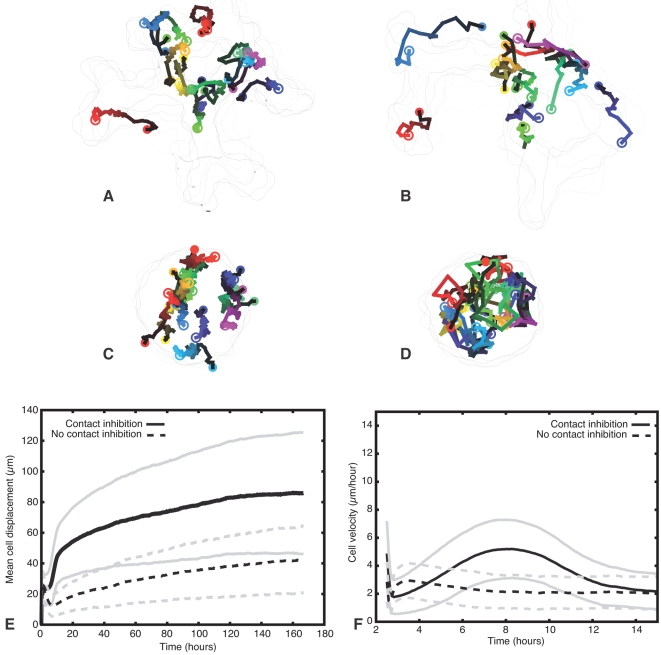
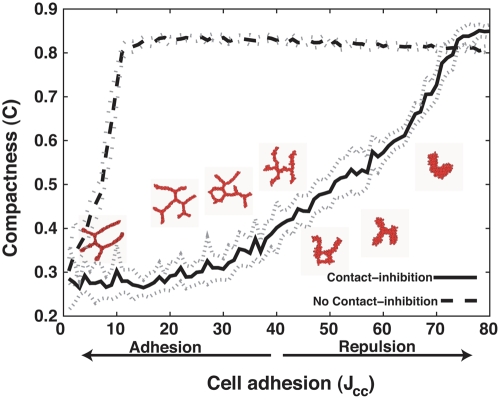
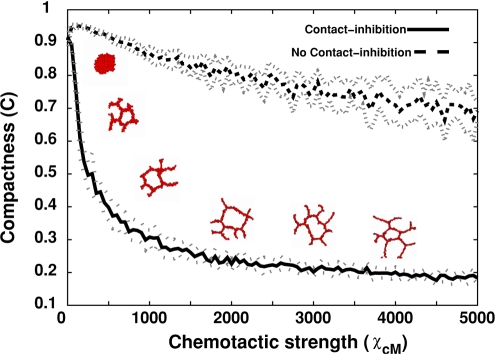
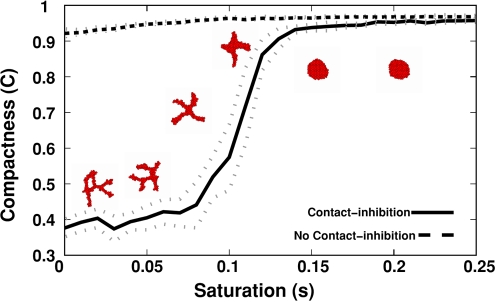
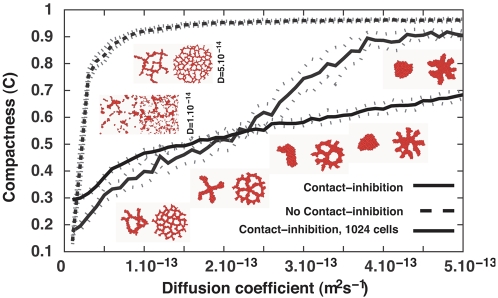
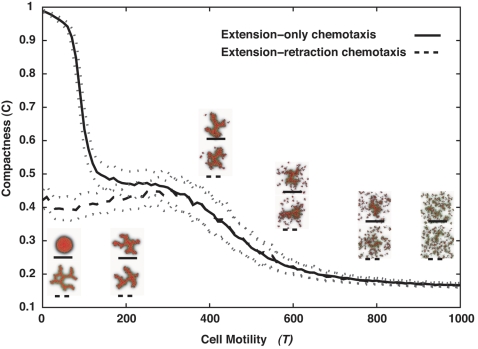
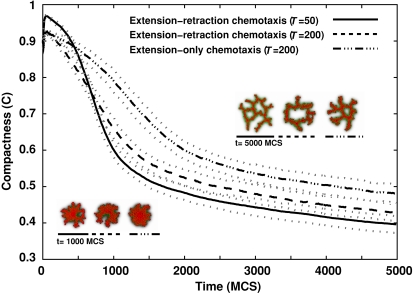
Blood vessels form either when dispersed endothelial cells (the cells lining the inner walls of fully formed blood vessels) organize into a vessel network (vasculogenesis), or by sprouting or splitting of existing blood vessels (angiogenesis). Although they are closely related biologically, no current model explains both phenomena with a single biophysical mechanism. Most computational models describe sprouting at the level of the blood vessel, ignoring how cell behavior drives branch splitting during sprouting. We present a cell-based, Glazier-Graner-Hogeweg model (also called Cellular Potts Model) simulation of the initial patterning before the vascular cords form lumens, based on plausible behaviors of endothelial cells. The endothelial cells secrete a chemoattractant, which attracts other endothelial cells. As in the classic Keller-Segel model, chemotaxis by itself causes cells to aggregate into isolated clusters. However, including experimentally observed VE-cadherin-mediated contact inhibition of chemotaxis in the simulation causes randomly distributed cells to organize into networks and cell aggregates to sprout, reproducing aspects of both de novo and sprouting blood-vessel growth. We discuss two branching instabilities responsible for our results. Cells at the surfaces of cell clusters attempting to migrate to the centers of the clusters produce a buckling instability. In a model variant that eliminates the surface-normal force, a dissipative mechanism drives sprouting, with the secreted chemical acting both as a chemoattractant and as an inhibitor of pseudopod extension. Both mechanisms would also apply if force transmission through the extracellular matrix rather than chemical signaling mediated cell-cell interactions. The branching instabilities responsible for our results, which result from contact inhibition of chemotaxis, are both generic developmental mechanisms and interesting examples of unusual patterning instabilities.
血管的形成方式有两种,一种是分散的内皮细胞(构成成熟血管内壁的细胞)组织成血管网络(血管发生),另一种是现有血管的出芽或分支(血管生成)。尽管它们在生物学上密切相关,但目前没有一个模型能用单一的生物物理机制来解释这两种现象。大多数计算模型是在血管层面描述出芽过程,忽略了细胞行为如何在出芽过程中驱动分支分裂。我们基于内皮细胞合理的行为,提出了一种基于细胞的Glazier-Graner-Hogeweg模型(也称为细胞Potts模型),用于模拟血管索形成管腔之前的初始模式。内皮细胞分泌一种趋化因子,吸引其他内皮细胞。与经典的Keller-Segel模型一样,趋化作用本身会使细胞聚集形成孤立的簇。然而,在模拟中纳入实验观察到的VE-钙黏蛋白介导的趋化接触抑制,会使随机分布的细胞组织成网络,细胞聚集体出芽,再现了从头开始和出芽血管生长的各个方面。我们讨论了导致我们结果的两种分支不稳定性。试图迁移到细胞簇中心的细胞簇表面的细胞会产生屈曲不稳定性。在一个消除表面法向力的模型变体中,一种耗散机制驱动出芽,分泌的化学物质既作为趋化因子,又作为伪足延伸的抑制剂。如果通过细胞外基质进行力传递而不是化学信号介导的细胞间相互作用,这两种机制也同样适用。导致我们结果的分支不稳定性,是由趋化接触抑制引起的,它们既是一般的发育机制,也是不寻常模式不稳定性的有趣例子。