Chromosome Stability Section, National Institute of Environmental Health Sciences, National Institutes of Health, Research Triangle Park, North Carolina, United States of America.
PLoS Genet. 2009 Sep;5(9):e1000656. doi: 10.1371/journal.pgen.1000656. Epub 2009 Sep 18.
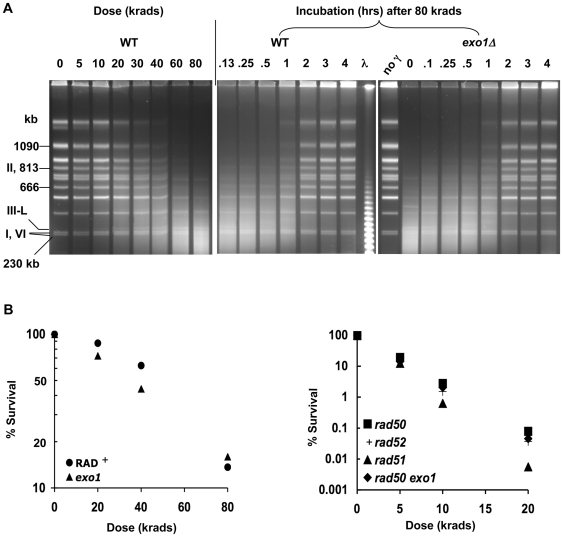
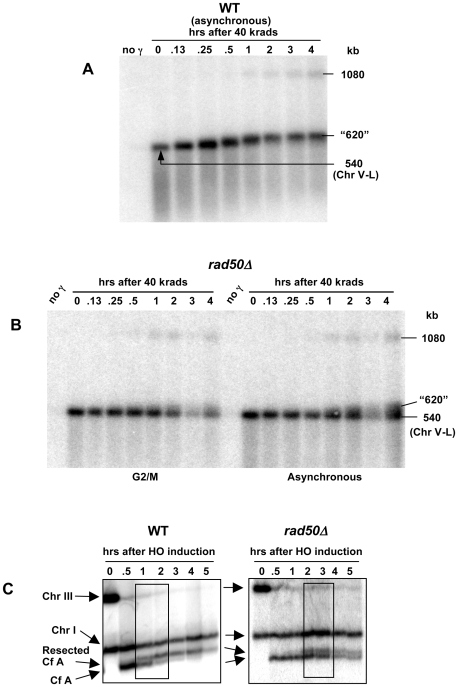
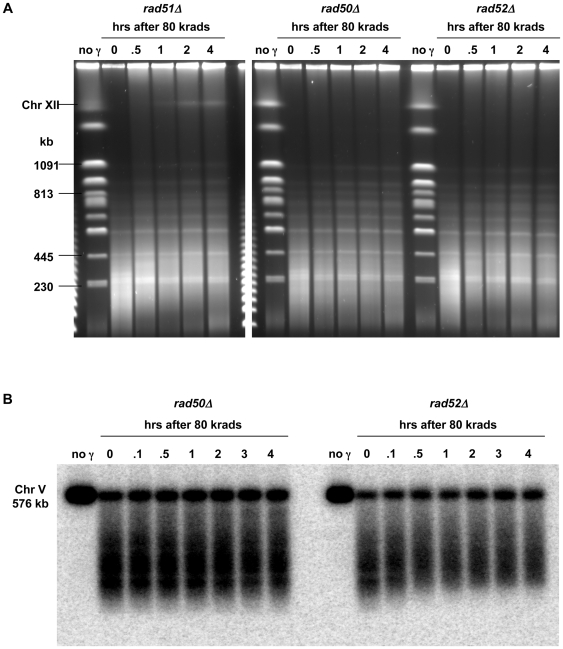
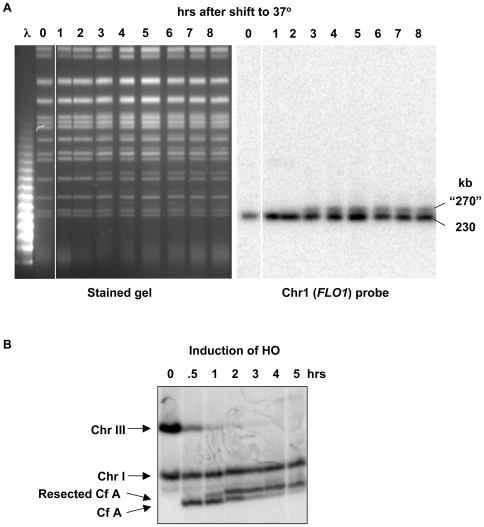
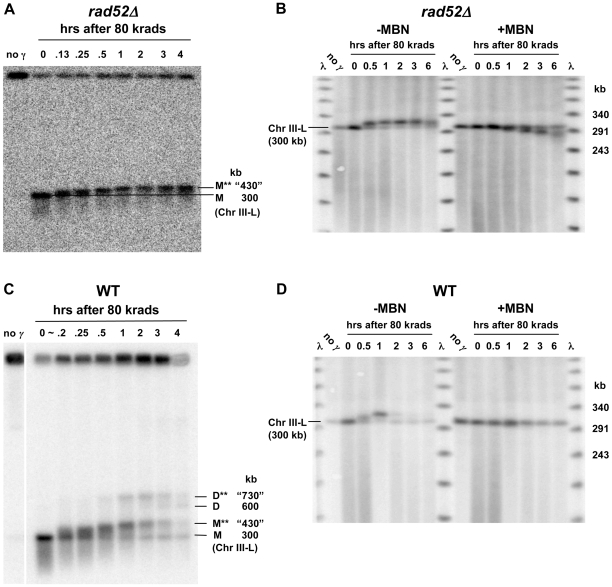
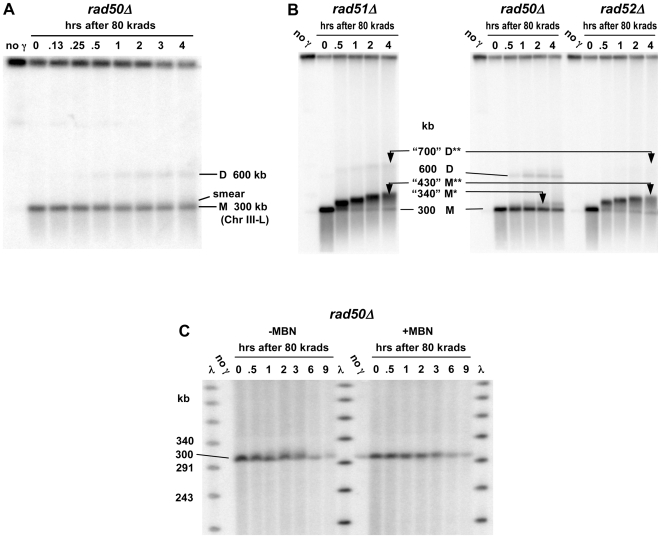
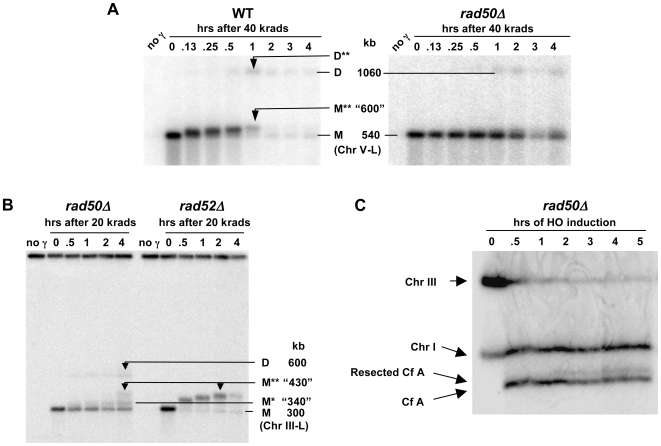
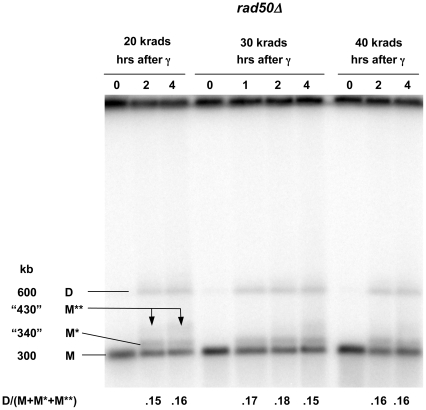
Resection of DNA double-strand break (DSB) ends is generally considered a critical determinant in pathways of DSB repair and genome stability. Unlike for enzymatically induced site-specific DSBs, little is known about processing of random "dirty-ended" DSBs created by DNA damaging agents such as ionizing radiation. Here we present a novel system for monitoring early events in the repair of random DSBs, based on our finding that single-strand tails generated by resection at the ends of large molecules in budding yeast decreases mobility during pulsed field gel electrophoresis (PFGE). We utilized this "PFGE-shift" to follow the fate of both ends of linear molecules generated by a single random DSB in circular chromosomes. Within 10 min after gamma-irradiation of G2/M arrested WT cells, there is a near-synchronous PFGE-shift of the linearized circular molecules, corresponding to resection of a few hundred bases. Resection at the radiation-induced DSBs continues so that by the time of significant repair of DSBs at 1 hr there is about 1-2 kb resection per DSB end. The PFGE-shift is comparable in WT and recombination-defective rad52 and rad51 strains but somewhat delayed in exo1 mutants. However, in rad50 and mre11 null mutants the initiation and generation of resected ends at radiation-induced DSB ends is greatly reduced in G2/M. Thus, the Rad50/Mre11/Xrs2 complex is responsible for rapid processing of most damaged ends into substrates that subsequently undergo recombinational repair. A similar requirement was found for RAD50 in asynchronously growing cells. Among the few molecules exhibiting shift in the rad50 mutant, the residual resection is consistent with resection at only one of the DSB ends. Surprisingly, within 1 hr after irradiation, double-length linear molecules are detected in the WT and rad50, but not in rad52, strains that are likely due to crossovers that are largely resection- and RAD50-independent.
DNA 双链断裂 (DSB) 末端的切除通常被认为是 DSB 修复和基因组稳定性途径中的关键决定因素。与酶诱导的特定位置 DSB 不同,对于诸如电离辐射等 DNA 损伤剂产生的随机“脏末端”DSB 的处理知之甚少。在这里,我们提出了一种监测随机 DSB 修复早期事件的新系统,该系统基于我们的发现,即通过在芽殖酵母中的大分子末端进行切除而产生的单链尾巴会降低脉冲场凝胶电泳 (PFGE) 期间的迁移率。我们利用这种“PFGE 迁移”来跟踪圆形染色体中单个随机 DSB 产生的线性分子的两个末端的命运。在 G2/M 期被γ射线照射的 WT 细胞 10 分钟内,线性化的圆形分子的 PFGE 迁移几乎是同步的,这对应于几百个碱基的切除。在辐射诱导的 DSB 处的切除继续进行,以至于在 1 小时时 DSB 修复有显著进展时,每个 DSB 末端大约有 1-2 kb 的切除。在 WT 和重组缺陷 rad52 和 rad51 菌株中,PFGE 迁移是可比的,但在 exo1 突变体中有些延迟。然而,在 rad50 和 mre11 缺失突变体中,在 G2/M 中,辐射诱导的 DSB 末端处的起始和产生的切除末端大大减少。因此,Rad50/Mre11/Xrs2 复合物负责将大多数受损末端快速加工成随后经历重组修复的底物。在同步生长的细胞中也发现 RAD50 具有类似的需求。在 rad50 突变体中表现出迁移的少数分子中,残留的切除与 DSB 末端之一的切除一致。令人惊讶的是,在照射后 1 小时内,在 WT 和 rad50 中检测到双长度的线性分子,但在 rad52 中没有,这可能是由于主要依赖于切除和 RAD50 不依赖的交叉。