Fundación Instituto Leloir - CONICET, Universidad de Buenos Aires, Argentina.
DNA Repair (Amst). 2010 Apr 4;9(4):358-64. doi: 10.1016/j.dnarep.2009.12.003. Epub 2010 Jan 8.
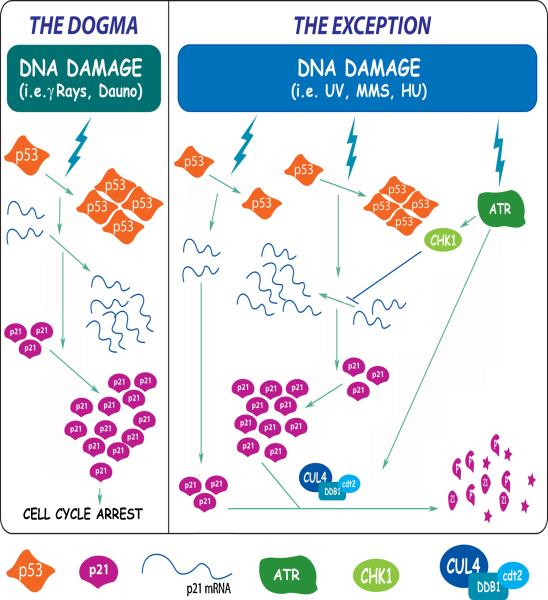
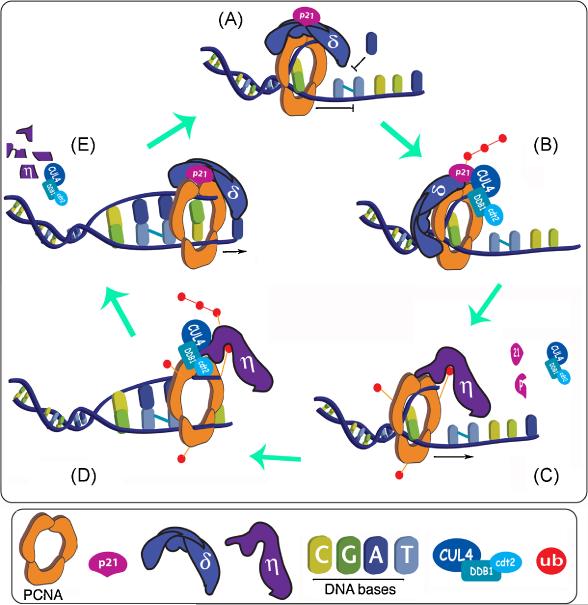
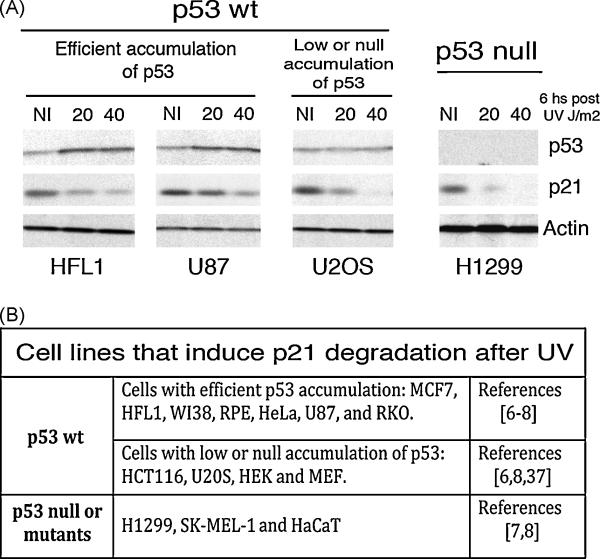
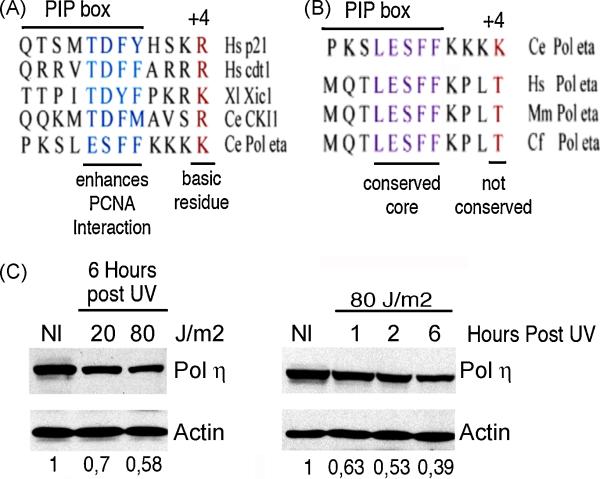
While many are the examples of DNA damaging treatments that induce p21 accumulation, the conception of p21 upregulation as the universal response to genotoxic stress has come to an end. Compelling evidences have demonstrated the existence of converging signals that negatively regulate p21 bellow basal levels when replication forks are blocked. Moreover, conclusive reports identified the E3-ligase CRL4(CDT2) (CUL4-DDB1-CDT2) as the enzymatic complex that promotes p21 proteolysis when treatments such as UV irradiation trigger replication fork stress. A pre-requisite for CRL4(CDT2)-driven proteolysis is the interaction of p21 with PCNA. Interestingly as well, CRL4(CDT2)-dependent proteolysis is not limited to p21 and affects other PCNA partners, including the specialized DNA polymerase eta (pol eta). These recent discoveries are particularly intriguing since the UV-induced degradation of p21 has been shown to be required for efficient pol eta recruitment to DNA lesions. Herein we review the findings that lead to the identification of the molecular mechanism that triggers damage-induced PCNA-coupled protein proteolysis. We propose a novel model in which CRL4(CDT2)-dependent protein degradation facilitates a sequential and dynamic exchange between PIP box bearing proteins at stall forks during Translesion DNA synthesis (TLS). Moreover, given the tight spatiotemporal control that CRL4(CDT2)-driven proteolysis is able to confer to PCNA-regulated processes, we discuss the impact that this degradation mechanism might have in other molecular switches associated with the repair of damaged DNA.
虽然有许多 DNA 损伤处理方法可诱导 p21 积累的例子,但 p21 上调作为对遗传毒性应激的普遍反应的概念已经结束。令人信服的证据表明,当复制叉受阻时,存在负调节 p21 低于基础水平的汇聚信号。此外,确凿的报告确定了 E3 连接酶 CRL4(CDT2)(CUL4-DDB1-CDT2)作为当诸如紫外线照射等处理引发复制叉应激时促进 p21 蛋白水解的酶复合物。CRL4(CDT2)驱动的蛋白水解的前提是 p21 与 PCNA 的相互作用。同样有趣的是,CRL4(CDT2)依赖性蛋白水解不仅限于 p21,还影响其他 PCNA 伴侣,包括专门的 DNA 聚合酶 eta(pol eta)。这些最近的发现特别有趣,因为已经表明紫外线诱导的 p21 降解对于有效的 pol eta 募集到 DNA 损伤是必需的。本文综述了导致鉴定触发损伤诱导的 PCNA 偶联蛋白蛋白水解的分子机制的发现。我们提出了一个新模型,其中 CRL4(CDT2)依赖性蛋白降解促进在跨损伤 DNA 合成(TLS)过程中停滞叉处带有 PIP 盒的蛋白之间的顺序和动态交换。此外,鉴于 CRL4(CDT2)驱动的蛋白水解能够赋予 PCNA 调节的过程的紧密时空控制,我们讨论了这种降解机制可能对与受损 DNA修复相关的其他分子开关的影响。