Department of Chemistry, Harding University, Searcy, Arkansas 72143, USA.
Biochemistry. 2010 Mar 16;49(10):2097-109. doi: 10.1021/bi901977k.
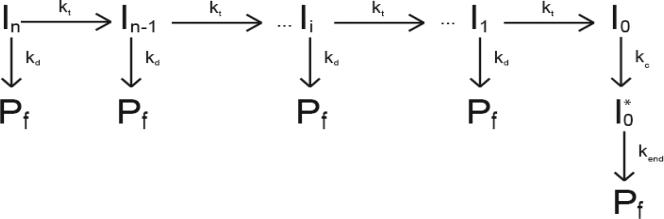
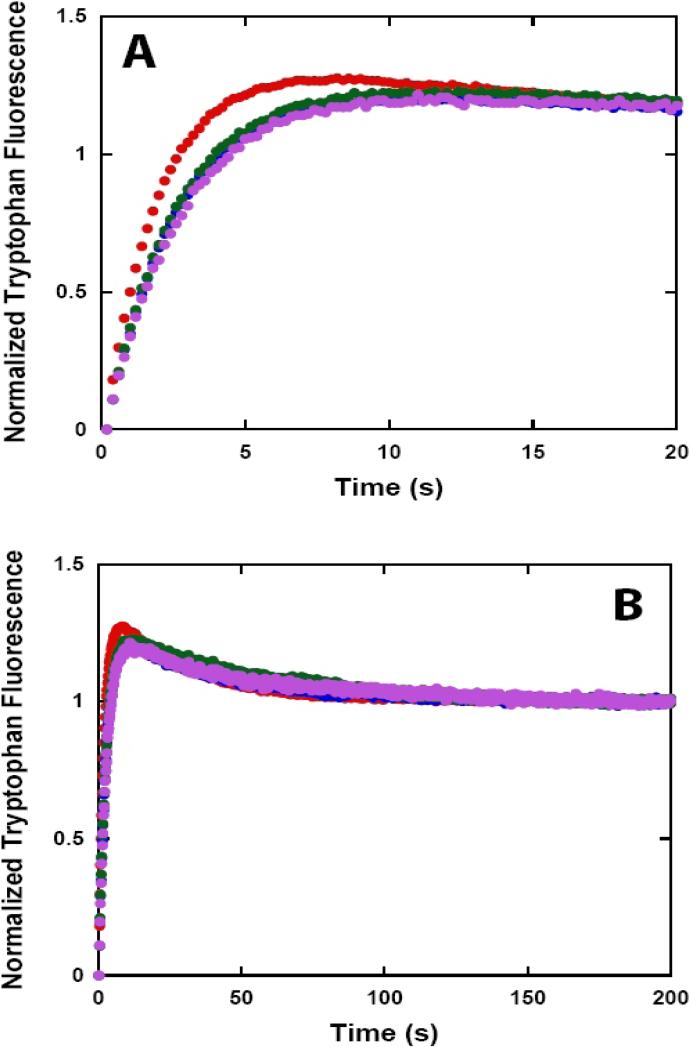
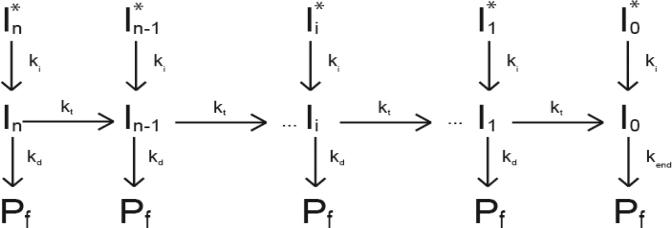
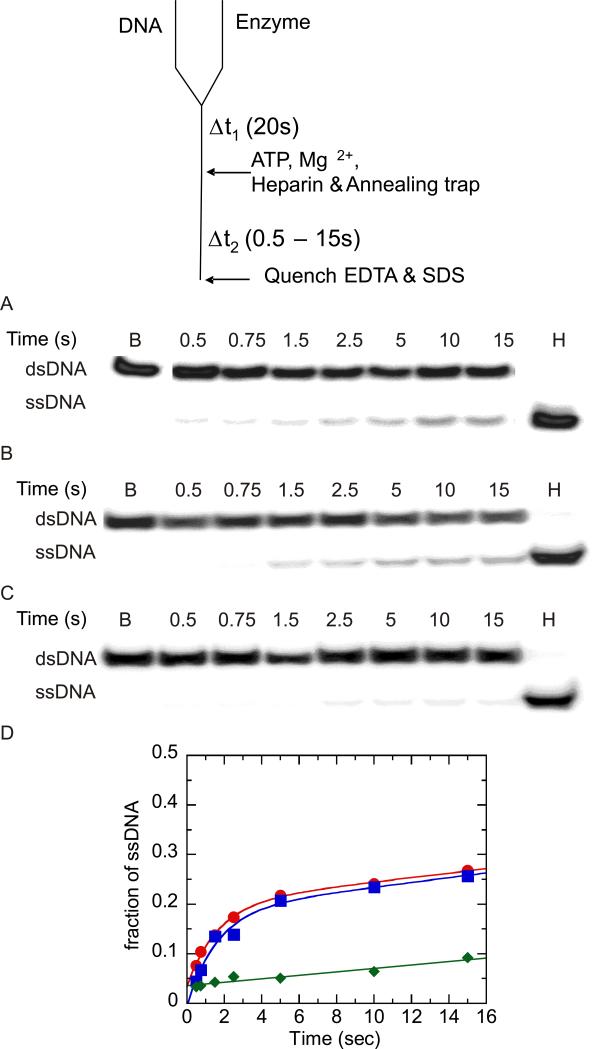
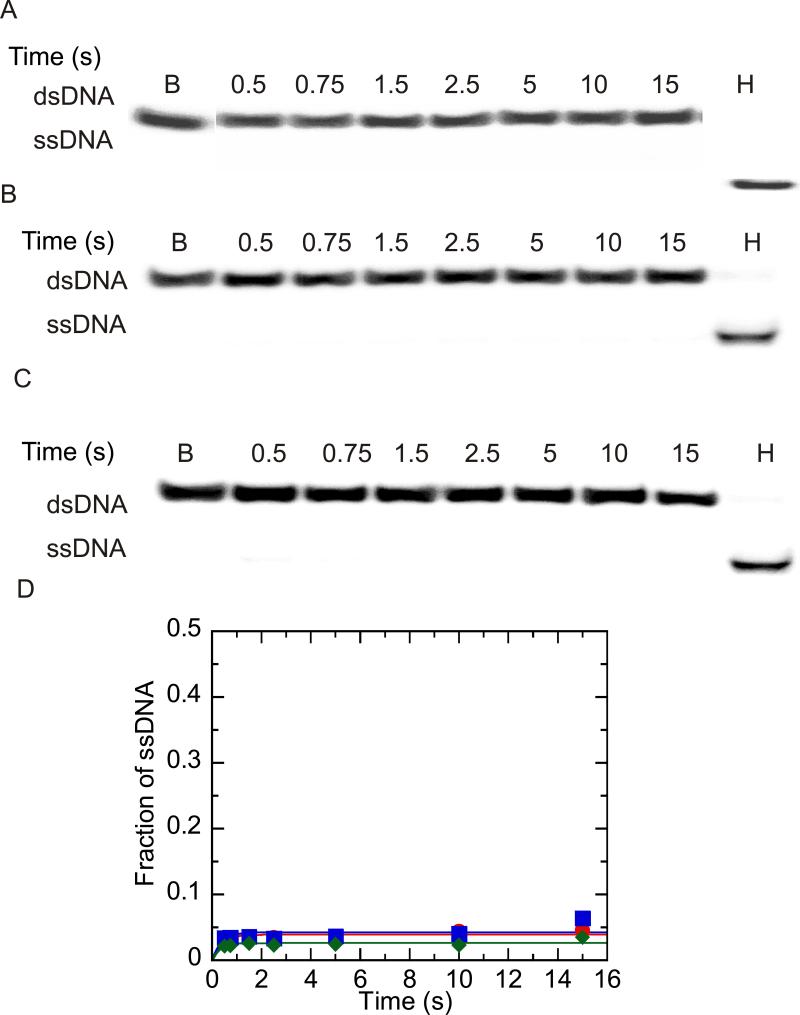
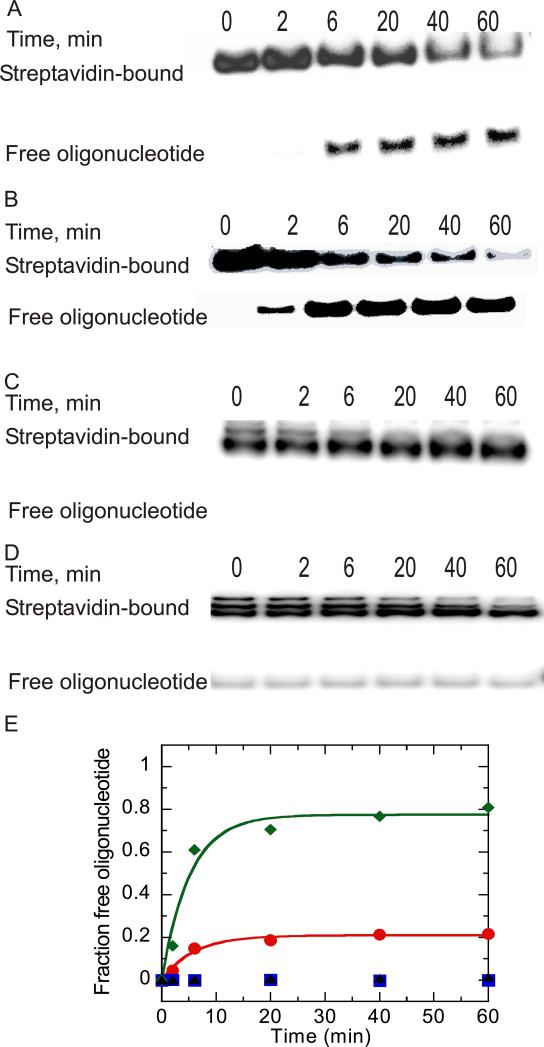
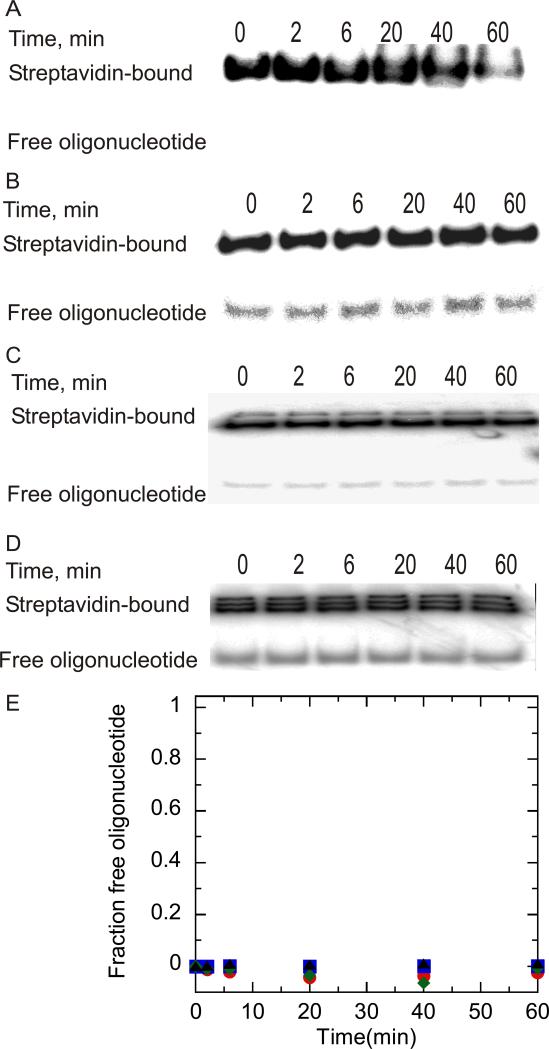
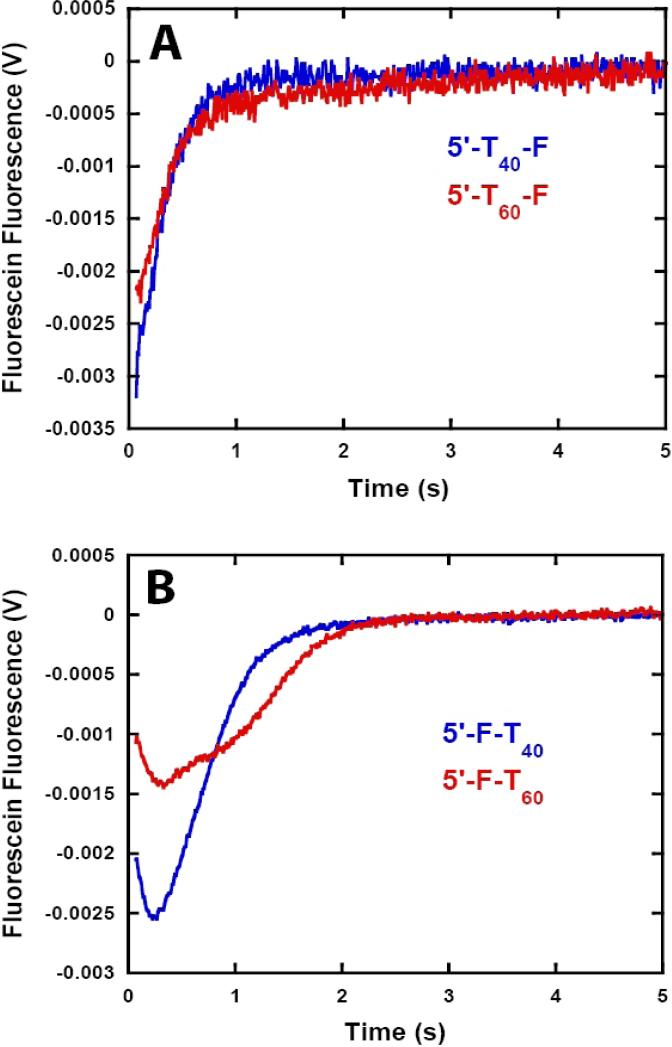
Helicases are motor proteins that are involved in DNA and RNA metabolism, replication, recombination, transcription, and repair. The motors are powered by ATP binding and hydrolysis. Hepatitis C virus encodes a helicase called nonstructural protein (NS3). NS3 possesses protease and helicase activities on its N-terminal and C-terminal domains, respectively. The helicase domain of NS3 is termed NS3h. In vitro, NS3h catalyzes RNA and DNA unwinding in a 3'-5' direction. The directionality of unwinding is thought to arise in part from the enzyme's ability to translocate along DNA, but translocation has not been shown explicitly. We examined the DNA translocase activity of NS3h by using single-stranded oligonucleotide substrates containing a fluorescent probe on the 5' end. NS3h can bind to the ssDNA and in the presence of ATP move toward the 5' end. When the enzyme encounters the fluorescent probe, a fluorescence change is observed that allows translocation to be characterized. Under conditions that favor binding of one NS3h per DNA substrate (100 nM NS3h and 200 nM oligonucleotide), we find that NS3h translocates on ssDNA at a rate of 46 +/- 5 nucleotides/s, and that it can move for 230 +/- 60 nucleotides before dissociating from the DNA. The translocase activity of some helicases is responsible for displacing proteins that are bound to DNA. We studied protein displacement by using a ssDNA oligonucleotide covalently linked to biotin on the 5' end. Upon addition of streptavidin, a "protein block" was placed in the pathway of the helicase. Interestingly, NS3h was unable to displace streptavidin from the end of the oligonucleotide, despite its ability to translocate along the DNA. The DNA unwinding activity of NS3h was examined using a 22 bp duplex DNA substrate under conditions that were identical to those used to study translocation. NS3h exhibited little or no DNA unwinding under single-cycle conditions, supporting the conclusion that NS3h is a relatively poor helicase in its monomeric form, as has been reported. In summary, NS3h translocates on ssDNA as a monomer, but the translocase activity does not correspond to comparable DNA unwinding activity or protein displacement activity under identical conditions.
解旋酶是参与 DNA 和 RNA 代谢、复制、重组、转录和修复的马达蛋白。这些马达蛋白通过 ATP 结合和水解来提供动力。丙型肝炎病毒编码一种称为非结构蛋白 (NS3) 的解旋酶。NS3 的 N 端和 C 端结构域分别具有蛋白酶和解旋酶活性。NS3 的解旋酶结构域称为 NS3h。在体外,NS3h 以 3'-5'方向催化 RNA 和 DNA 的解旋。解旋的方向性部分源于酶沿 DNA 易位的能力,但尚未明确显示易位。我们使用含有 5'端荧光探针的单链寡核苷酸底物来检查 NS3h 的 DNA 转位酶活性。NS3h 可以与 ssDNA 结合,并在 ATP 的存在下向 5'端移动。当酶遇到荧光探针时,观察到荧光变化,从而可以对转位进行表征。在有利于每个 DNA 底物结合一个 NS3h 的条件下(100 nM NS3h 和 200 nM 寡核苷酸),我们发现 NS3h 在 ssDNA 上的转位速度为 46 +/- 5 个核苷酸/s,并且可以在从 DNA 上解离之前移动 230 +/- 60 个核苷酸。一些解旋酶的转位酶活性负责置换与 DNA 结合的蛋白质。我们使用 5'端通过共价键连接生物素的 ssDNA 寡核苷酸研究蛋白质置换。加入链霉亲和素后,在解旋酶的路径上放置了一个“蛋白质块”。有趣的是,尽管 NS3h 能够沿 DNA 易位,但它无法从寡核苷酸的末端置换链霉亲和素。在与研究转位相同的条件下,使用 22 个碱基对的双链 DNA 底物研究了 NS3h 的 DNA 解旋活性。在单循环条件下,NS3h 几乎没有或没有 DNA 解旋,这支持了 NS3h 在单体形式下是一种相对较差的解旋酶的结论,正如已经报道的那样。总之,NS3h 作为单体在 ssDNA 上易位,但在相同条件下,转位酶活性与可比的 DNA 解旋活性或蛋白质置换活性不对应。