Department of Medical Parasitology and Infection Biology, Swiss Tropical Institute, University of Basel, Basel, Switzerland.
PLoS Pathog. 2010 Feb 26;6(2):e1000784. doi: 10.1371/journal.ppat.1000784.
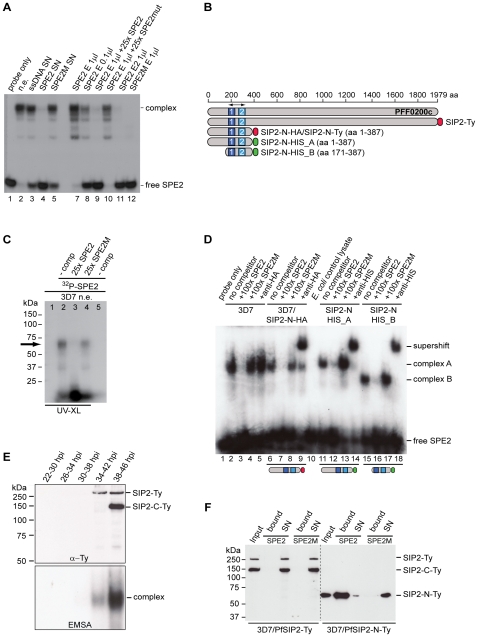
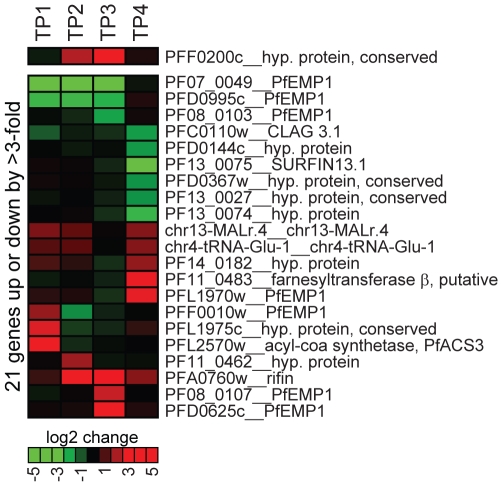
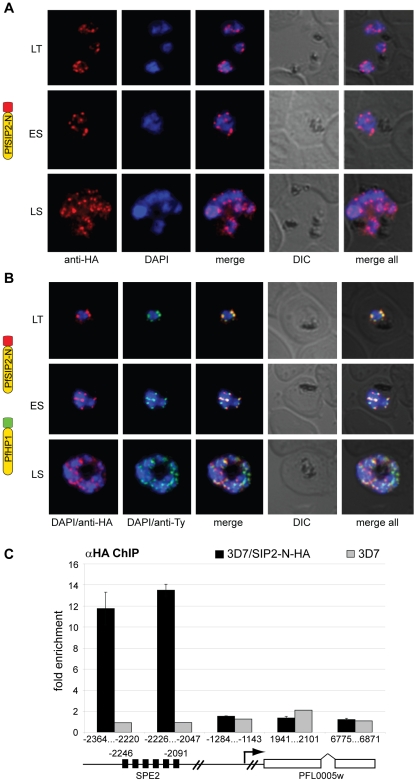
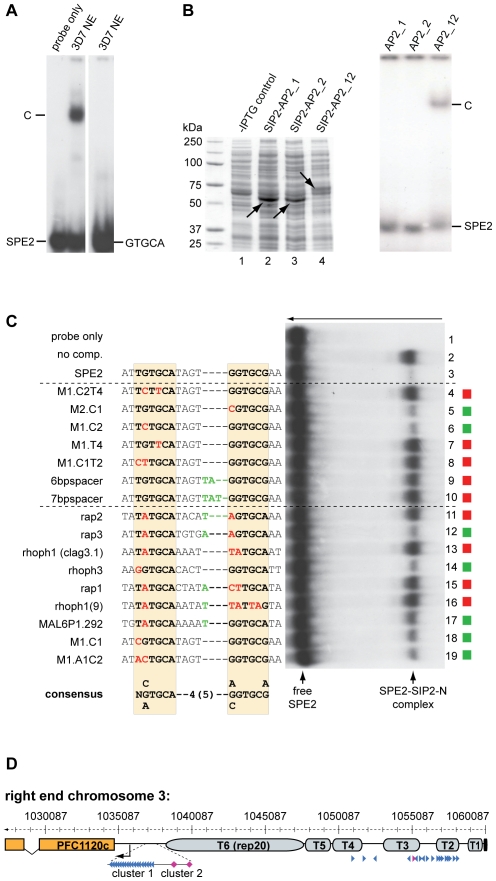
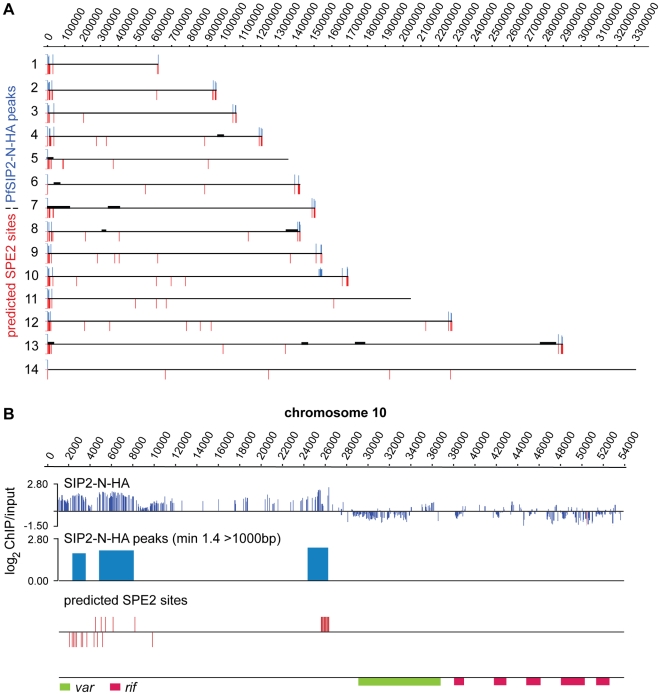
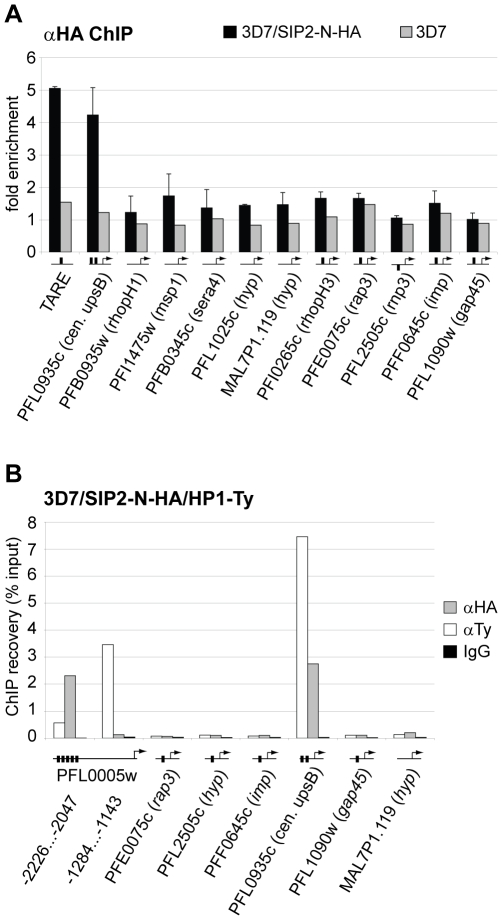
The heterochromatic environment and physical clustering of chromosome ends at the nuclear periphery provide a functional and structural framework for antigenic variation and evolution of subtelomeric virulence gene families in the malaria parasite Plasmodium falciparum. While recent studies assigned important roles for reversible histone modifications, silent information regulator 2 and heterochromatin protein 1 (PfHP1) in epigenetic control of variegated expression, factors involved in the recruitment and organization of subtelomeric heterochromatin remain unknown. Here, we describe the purification and characterization of PfSIP2, a member of the ApiAP2 family of putative transcription factors, as the unknown nuclear factor interacting specifically with cis-acting SPE2 motif arrays in subtelomeric domains. Interestingly, SPE2 is not bound by the full-length protein but rather by a 60kDa N-terminal domain, PfSIP2-N, which is released during schizogony. Our experimental re-definition of the SPE2/PfSIP2-N interaction highlights the strict requirement of both adjacent AP2 domains and a conserved bipartite SPE2 consensus motif for high-affinity binding. Genome-wide in silico mapping identified 777 putative binding sites, 94% of which cluster in heterochromatic domains upstream of subtelomeric var genes and in telomere-associated repeat elements. Immunofluorescence and chromatin immunoprecipitation (ChIP) assays revealed co-localization of PfSIP2-N with PfHP1 at chromosome ends. Genome-wide ChIP demonstrated the exclusive binding of PfSIP2-N to subtelomeric SPE2 landmarks in vivo but not to single chromosome-internal sites. Consistent with this specialized distribution pattern, PfSIP2-N over-expression has no effect on global gene transcription. Hence, contrary to the previously proposed role for this factor in gene activation, our results provide strong evidence for the first time for the involvement of an ApiAP2 factor in heterochromatin formation and genome integrity. These findings are highly relevant for our understanding of chromosome end biology and variegated expression in P. falciparum and other eukaryotes, and for the future analysis of the role of ApiAP2-DNA interactions in parasite biology.
异染色质环境和染色体末端在核周的物理聚集为疟原虫 Plasmodium falciparum 中的抗原变异和端粒病毒基因家族的进化提供了功能和结构框架。虽然最近的研究将可逆组蛋白修饰、沉默信息调节因子 2 和异染色质蛋白 1 (PfHP1) 赋予了表观遗传控制变异性表达的重要作用,但参与端粒异染色质募集和组织的因素仍不清楚。在这里,我们描述了 PfSIP2 的纯化和表征,PfSIP2 是假定转录因子 ApiAP2 家族的成员,是与端粒域中顺式作用 SPE2 基序阵列特异性相互作用的未知核因子。有趣的是,SPE2 不是由全长蛋白结合,而是由在裂殖体期间释放的 60kDa N 端结构域 PfSIP2-N 结合。我们对 SPE2/PfSIP2-N 相互作用的实验重新定义突出了两个相邻 AP2 结构域和保守的二分 SPE2 共识基序对于高亲和力结合的严格要求。全基因组计算机绘图鉴定了 777 个推定的结合位点,其中 94%聚集在端粒相关重复元件和端粒下游的异染色质结构域中。免疫荧光和染色质免疫沉淀 (ChIP) 实验显示 PfSIP2-N 与 PfHP1 在染色体末端共定位。全基因组 ChIP 表明 PfSIP2-N 仅在体内与端粒特异性 SPE2 地标结合,而不与单个染色体内部位点结合。与这种特殊的分布模式一致,PfSIP2-N 的过表达对全局基因转录没有影响。因此,与该因子在基因激活中的先前提出的作用相反,我们的结果首次提供了强有力的证据,证明 ApiAP2 因子参与异染色质形成和基因组完整性。这些发现对于我们理解疟原虫和其他真核生物中的染色体末端生物学和变异性表达以及未来分析 ApiAP2-DNA 相互作用在寄生虫生物学中的作用具有重要意义。