den Braber Anouk, van 't Ent Dennis, Cath Danielle C, Wagner Judith, Boomsma Dorret I, de Geus Eco J C
Department of Biological Psychology, VU University, van der Boechorststraat 1, Amsterdam, The Netherlands.
Brain. 2010 Oct;133(10):3123-40. doi: 10.1093/brain/awq229. Epub 2010 Sep 7.
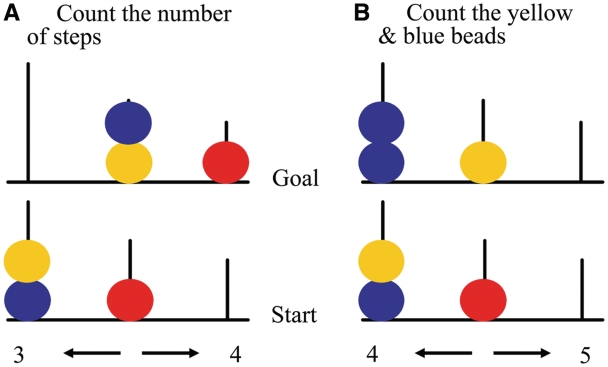
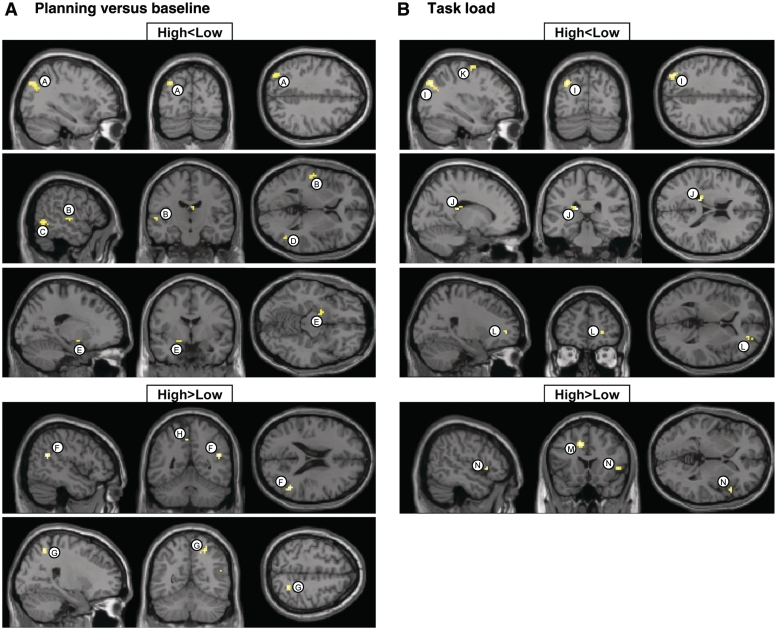
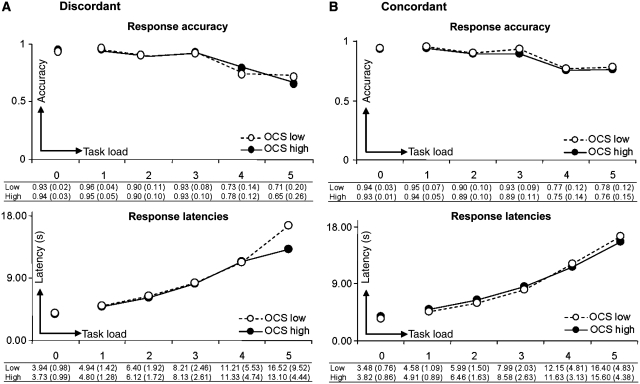
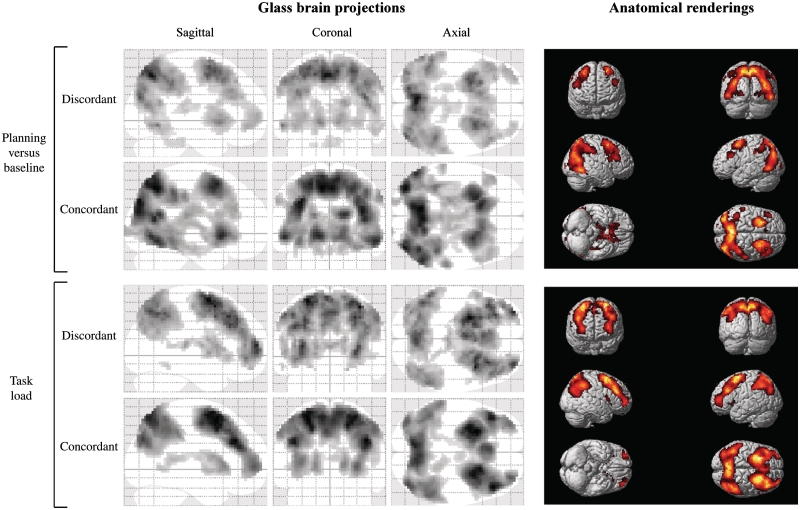
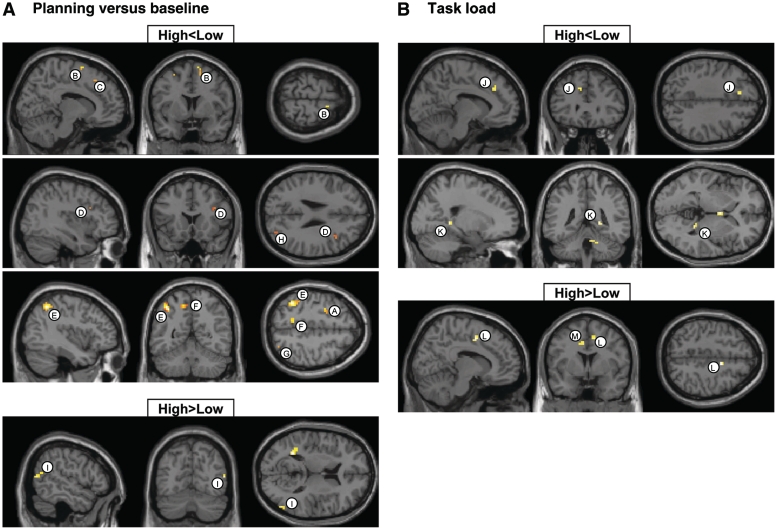
Neuroimaging studies have indicated abnormalities in cortico-striatal-thalamo-cortical circuits in patients with obsessive-compulsive disorder compared with controls. However, there are inconsistencies between studies regarding the exact set of brain structures involved and the direction of anatomical and functional changes. These inconsistencies may reflect the differential impact of environmental and genetic risk factors for obsessive-compulsive disorder on different parts of the brain. To distinguish between functional brain changes underlying environmentally and genetically mediated obsessive-compulsive disorder, we compared task performance and brain activation during a Tower of London planning paradigm in monozygotic twins discordant (n=38) or concordant (n=100) for obsessive-compulsive symptoms. Twins who score high on obsessive-compulsive symptoms can be considered at high risk for obsessive-compulsive disorder. We found that subjects at high risk for obsessive-compulsive disorder did not differ from the low-risk subjects behaviourally, but we obtained evidence that the high-risk subjects differed from the low-risk subjects in the patterns of brain activation accompanying task execution. These regions can be separated into those that were affected by mainly environmental risk (dorsolateral prefrontal cortex and lingual cortex), genetic risk (frontopolar cortex, inferior frontal cortex, globus pallidus and caudate nucleus) and regions affected by both environmental and genetic risk factors (cingulate cortex, premotor cortex and parts of the parietal cortex). Our results suggest that neurobiological changes related to obsessive-compulsive symptoms induced by environmental factors involve primarily the dorsolateral prefrontal cortex, whereas neurobiological changes induced by genetic factors involve orbitofrontal-basal ganglia structures. Regions showing similar changes in high-risk twins from discordant and concordant pairs may be part of compensatory networks that keep planning performance intact, in spite of cortico-striatal-thalamo-cortical deficits.
神经影像学研究表明,与对照组相比,强迫症患者的皮质-纹状体-丘脑-皮质回路存在异常。然而,关于具体涉及的脑结构集以及解剖和功能变化的方向,不同研究之间存在不一致之处。这些不一致可能反映了强迫症的环境和遗传风险因素对大脑不同部位的不同影响。为了区分环境介导和遗传介导的强迫症背后的大脑功能变化,我们比较了在伦敦塔计划范式中,同卵双胞胎中患强迫症症状不一致(n = 38)或一致(n = 100)的双胞胎在任务表现和大脑激活方面的差异。在强迫症症状上得分高的双胞胎可被视为患强迫症的高风险人群。我们发现,患强迫症高风险的受试者在行为上与低风险受试者没有差异,但我们获得的证据表明,高风险受试者与低风险受试者在任务执行时的大脑激活模式上存在差异。这些区域可分为主要受环境风险影响的区域(背外侧前额叶皮质和舌回皮质)、遗传风险影响的区域(额极皮质、额下回皮质、苍白球和尾状核)以及受环境和遗传风险因素共同影响的区域(扣带回皮质、运动前皮质和部分顶叶皮质)。我们的结果表明,环境因素诱发的与强迫症症状相关的神经生物学变化主要涉及背外侧前额叶皮质,而遗传因素诱发的神经生物学变化涉及眶额-基底神经节结构。在不一致和一致的双胞胎对中,高风险双胞胎中显示出类似变化的区域可能是补偿网络的一部分,尽管存在皮质-纹状体-丘脑-皮质缺陷,但仍能保持计划表现完好无损。