Department of Cell Biology, Harvard Medical School, 240 Longwood Avenue, Boston, MA 02115, USA.
Mol Cell. 2010 Nov 24;40(4):671-81. doi: 10.1016/j.molcel.2010.11.002.
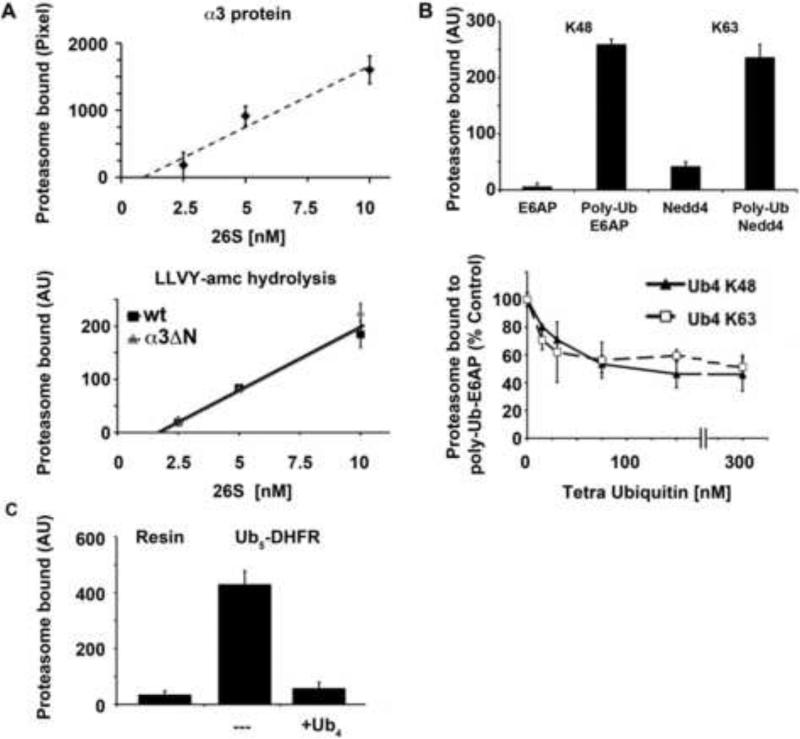
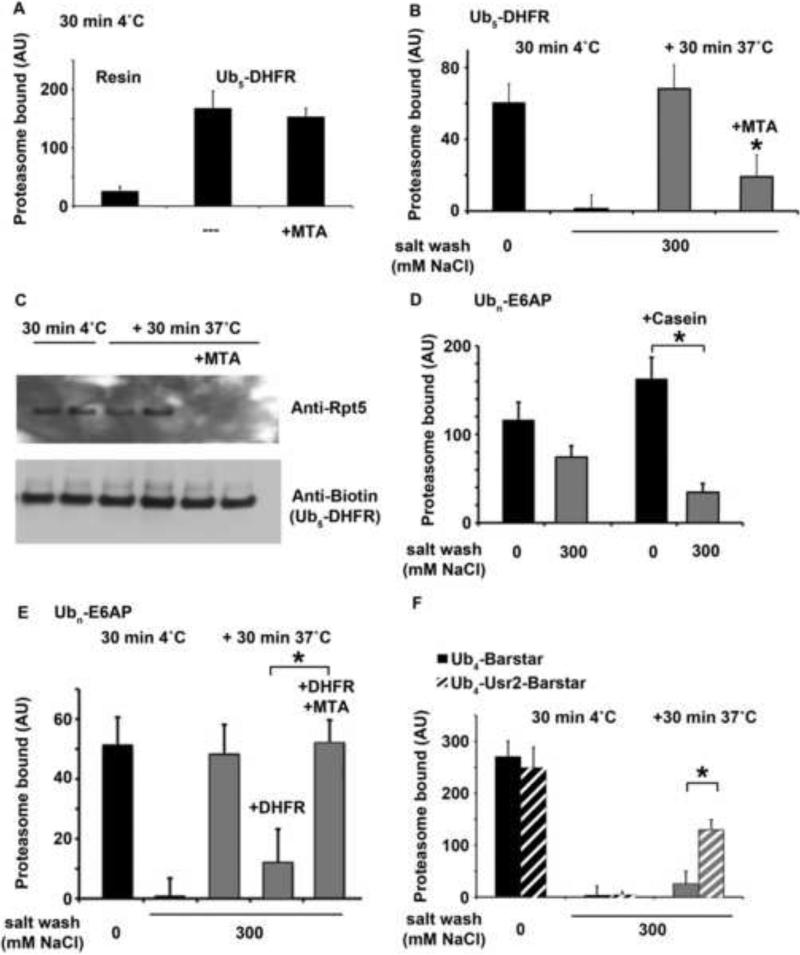
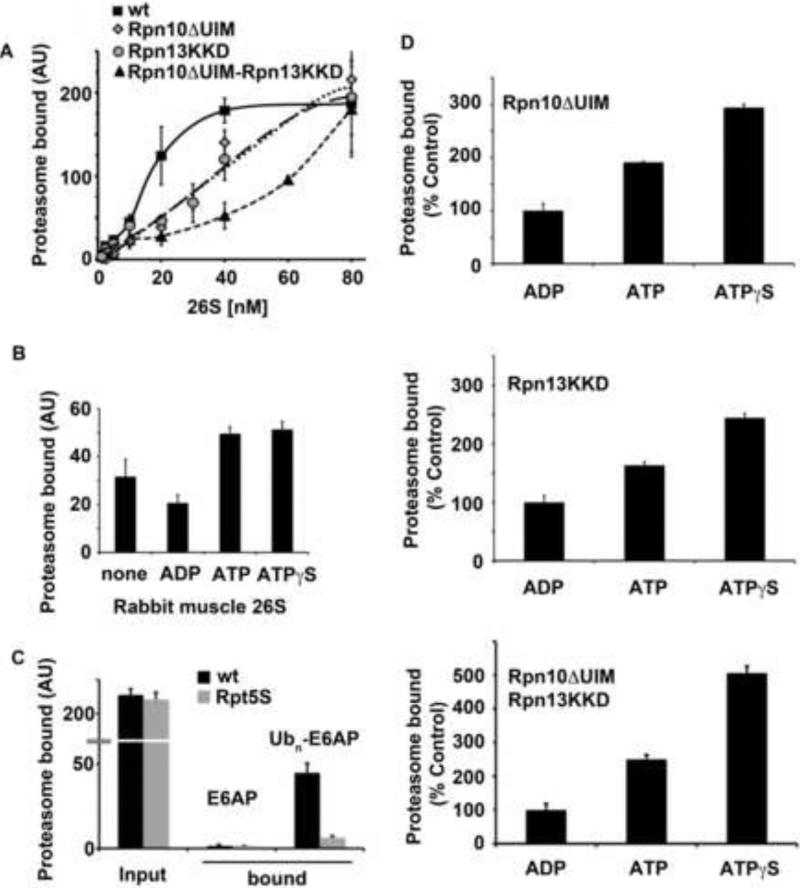
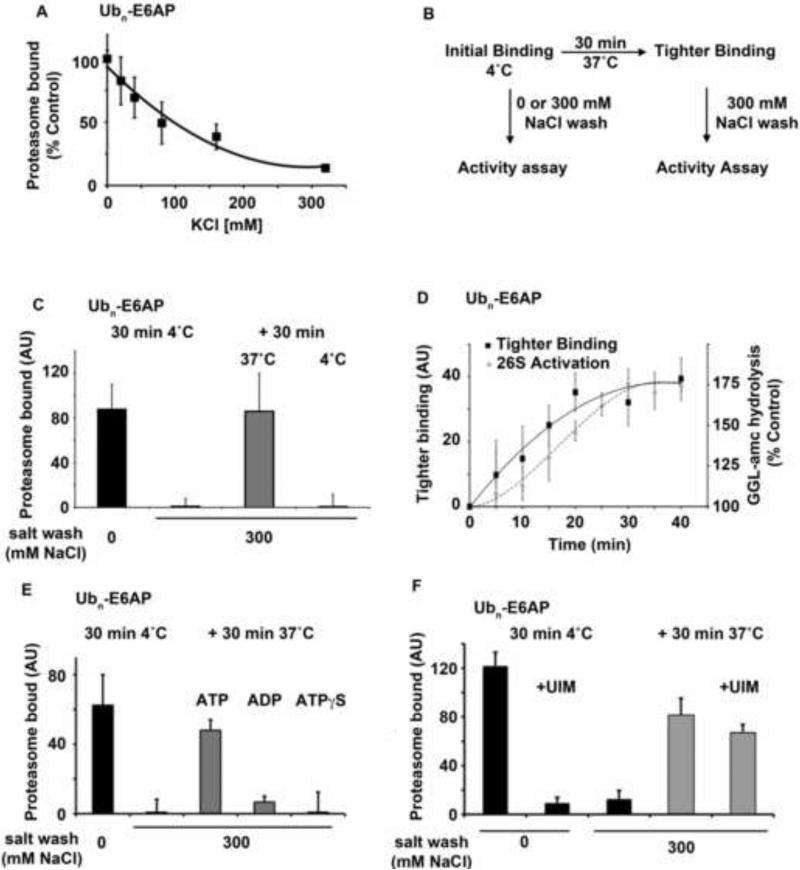
Eukaryotic cells target proteins for degradation by the 26S proteasome by attaching a ubiquitin chain. Using a rapid assay, we analyzed the initial binding of ubiquitinated proteins to purified 26S particles as an isolated process at 4°C. Subunits Rpn10 and Rpn13 contribute equally to the high-affinity binding of ubiquitin chains, but in their absence, ubiquitin conjugates bind to another site with 4-fold lower affinity. Conjugate binding is stimulated 2- to 4-fold by binding of ATP or the nonhydrolyzable analog, ATPγS (but not ADP), to the 19S ATPases. Following this initial, reversible association, ubiquitin conjugates at 37°C become more tightly bound through a step that requires ATP hydrolysis and a loosely folded domain on the protein, but appears independent of ubiquitin. Unfolded or loosely folded polypeptides can inhibit this tighter binding. This commitment step precedes substrate deubiquitination and allows for selection of ubiquitinated proteins capable of being unfolded and efficiently degraded.
真核细胞通过附着泛素链将蛋白质靶向 26S 蛋白酶体进行降解。我们使用快速测定法,在 4°C 下作为一个独立的过程分析了已泛素化的蛋白质与纯化的 26S 颗粒的初始结合。亚基 Rpn10 和 Rpn13 对泛素链的高亲和力结合贡献相等,但在它们缺失的情况下,泛素缀合物以低 4 倍亲和力结合到另一个位点。结合物结合通过结合到 19S ATP 酶的 ATP 或非水解类似物 ATPγS(但不是 ADP)而被刺激 2 至 4 倍。在这个初始的可逆结合之后,泛素缀合物在 37°C 下通过需要 ATP 水解和蛋白质上的松散折叠结构域的步骤更紧密地结合,但似乎与泛素无关。未折叠或松散折叠的多肽可以抑制这种更紧密的结合。此承诺步骤先于底物去泛素化,并且允许选择能够展开和有效降解的泛素化蛋白质。