Department of Pharmacology, College of Medicine, The University of Tennessee Health Science Center, Memphis, TN 38163, USA.
J Gen Physiol. 2011 Jan;137(1):93-110. doi: 10.1085/jgp.201010519. Epub 2010 Dec 13.
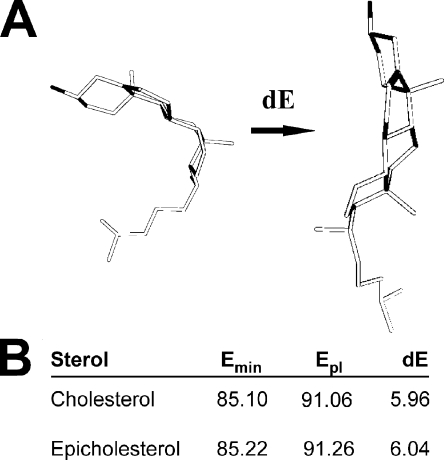
The activity (Po) of large-conductance voltage/Ca(2+)-gated K(+) (BK) channels is blunted by cholesterol levels within the range found in natural membranes. We probed BK channel-forming α (cbv1) subunits in phospholipid bilayers with cholesterol and related monohydroxysterols and performed computational dynamics to pinpoint the structural requirements for monohydroxysterols to reduce BK Po and obtain insights into cholesterol's mechanism of action. Cholesterol, cholestanol, and coprostanol reduced Po by shortening mean open and lengthening mean closed times, whereas epicholesterol, epicholestanol, epicoprostanol, and cholesterol trisnorcholenic acid were ineffective. Thus, channel inhibition by monohydroxysterols requires the β configuration of the C3 hydroxyl and is favored by the hydrophobic nature of the side chain, while having lax requirements on the sterol A/B ring fusion. Destabilization of BK channel open state(s) has been previously interpreted as reflecting increased bilayer lateral stress by cholesterol. Lateral stress is controlled by the sterol molecular area and lipid monolayer lateral tension, the latter being related to the sterol ability to adopt a planar conformation in lipid media. However, we found that the differential efficacies of monohydroxysterols to reduce Po (cholesterol≥coprostanol≥cholestanol>>>epicholesterol) did not follow molecular area rank (coprostanol>>epicholesterol>cholesterol>cholestanol). In addition, computationally predicted energies for cholesterol (effective BK inhibitor) and epicholesterol (ineffective) to adopt a planar conformation were similar. Finally, cholesterol and coprostanol reduced Po, yet these sterols have opposite effects on tight lipid packing and, likely, on lateral stress. Collectively, these findings suggest that an increase in bilayer lateral stress is unlikely to underlie the differential ability of cholesterol and related steroids to inhibit BK channels. Remarkably, ent-cholesterol (cholesterol mirror image) failed to reduce Po, indicating that cholesterol efficacy requires sterol stereospecific recognition by a protein surface. The BK channel phenotype resembled that of α homotetramers. Thus, we hypothesize that a cholesterol-recognizing protein surface resides at the BK α subunit itself.
大电导电压/钙(Ca2+)门控钾(K+)(BK)通道的活性(Po)在天然膜中发现的胆固醇水平范围内受到抑制。我们用胆固醇和相关的单羟基固醇探测 BK 通道形成的α(cbv1)亚基在磷脂双层中,并进行计算动力学,以确定单羟基固醇降低 BK Po 的结构要求,并深入了解胆固醇的作用机制。胆固醇、胆甾醇和粪甾醇通过缩短平均开放时间和延长平均关闭时间来降低 Po,而表胆固醇、表胆甾醇、表粪甾醇和胆固醇三降胆甾醇则无效。因此,单羟基固醇对通道的抑制作用需要 C3 羟基的β构型,并受到侧链疏水性的有利影响,而对甾醇 A/B 环融合的要求则较宽松。BK 通道开放状态的不稳定性先前被解释为反映胆固醇增加了双层侧向应力。侧向应力由甾醇分子面积和单层脂质侧向张力控制,后者与甾醇在脂质介质中采用平面构象的能力有关。然而,我们发现单羟基固醇降低 Po 的功效差异(胆固醇≥粪甾醇≥胆甾醇>>>表胆固醇)并没有遵循分子面积等级(粪甾醇>表胆固醇>胆固醇>胆甾醇)。此外,计算预测胆固醇(有效 BK 抑制剂)和表胆固醇(无效)采用平面构象的能量相似。最后,胆固醇和粪甾醇降低了 Po,但这些甾醇对紧密脂质堆积有相反的影响,可能对侧向应力也有影响。总的来说,这些发现表明,增加双层侧向应力不太可能是胆固醇和相关类固醇抑制 BK 通道的不同能力的基础。值得注意的是,对映胆固醇(胆固醇镜像)未能降低 Po,表明胆固醇的功效需要甾醇在蛋白质表面的立体特异性识别。BK 通道表型与α同四聚体相似。因此,我们假设胆固醇识别蛋白表面位于 BKα亚基本身。