Centre of Systems Biology at Edinburgh, University of Edinburgh, Edinburgh, Midlothian EH9 3JD, UK.
Curr Biol. 2011 Jan 25;21(2):120-5. doi: 10.1016/j.cub.2010.12.013. Epub 2011 Jan 13.
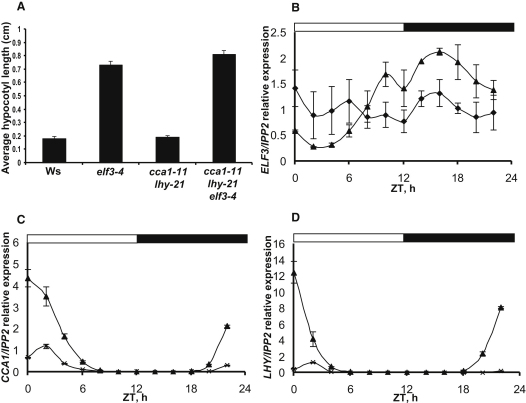
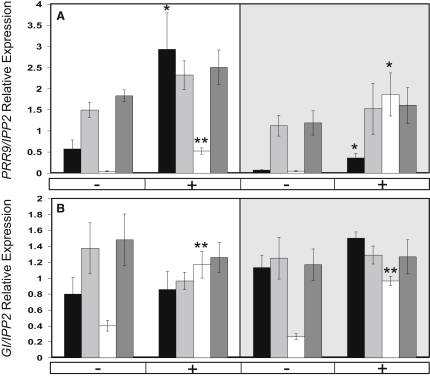
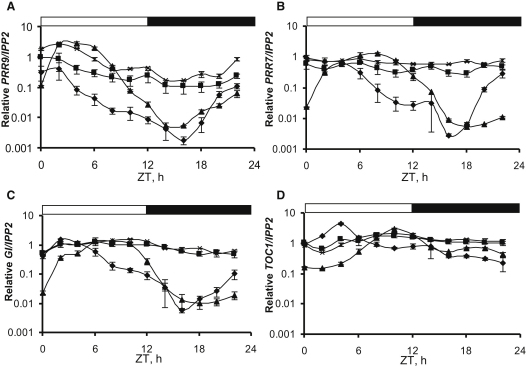
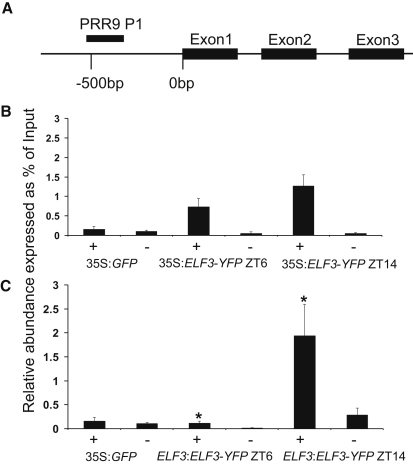
The circadian clock provides robust, ∼24 hr biological rhythms throughout the eukaryotes. The clock gene circuit in plants comprises interlocking transcriptional feedback loops, reviewed in [1], whereby the morning-expressed transcription factors CIRCADIAN CLOCK-ASSOCIATED 1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) repress the expression of evening genes, notably TIMING OF CAB EXPRESSION 1 (TOC1). EARLY FLOWERING 3 (ELF3) has been implicated as a repressor of light signaling to the clock [2, 3] and, paradoxically, as an activator of the light-induced genes CCA1 and LHY [4, 5]. We use cca1-11 lhy-21 elf3-4 plants to separate the repressive function of ELF3 from its downstream targets CCA1 and LHY. We further demonstrate that ELF3 associates physically with the promoter of PSEUDO-RESPONSE REGULATOR 9 (PRR9), a repressor of CCA1 and LHY expression, in a time-dependent fashion. The repressive function of ELF3 is thus consistent with indirect activation of LHY and CCA1, in a double-negative connection via a direct ELF3 target, PRR9. This mechanism reconciles the functions of ELF3 in the clock network during the night and points to further effects of ELF3 during the day.
生物钟为真核生物提供了强大的、近似 24 小时的生物节律。[1]综述了植物中的生物钟基因电路,包括互锁的转录反馈环,其中,在早晨表达的转录因子circadian clock-associated 1 (CCA1)和late elongated hypocotyl (LHY)抑制了 evening 基因的表达,尤其是 timing of CAB expression 1 (TOC1)。EARLY FLOWERING 3 (ELF3) 被认为是光信号到生物钟的抑制剂[2,3],但矛盾的是,它也是光诱导基因 CCA1 和 LHY 的激活剂[4,5]。我们使用 cca1-11 lhy-21 elf3-4 植物来分离 ELF3 的抑制功能与其下游靶基因 CCA1 和 LHY。我们进一步证明,ELF3 以时间依赖的方式与 PSEUDO-RESPONSE REGULATOR 9 (PRR9)的启动子结合,PRR9 是 CCA1 和 LHY 表达的抑制剂。因此,ELF3 的抑制功能与 LHY 和 CCA1 的间接激活一致,通过直接的 ELF3 靶标 PRR9 形成双负连接。该机制协调了 ELF3 在夜间生物钟网络中的功能,并指出了 ELF3 在白天的进一步作用。