Jelinek Ivett, Leonard Joshua N, Price Graeme E, Brown Kevin N, Meyer-Manlapat Anna, Goldsmith Paul K, Wang Yan, Venzon David, Epstein Suzanne L, Segal David M
Experimental Immunology Branch, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Bethesda, MD 20892, USA.
J Immunol. 2011 Feb 15;186(4):2422-9. doi: 10.4049/jimmunol.1002845. Epub 2011 Jan 17.
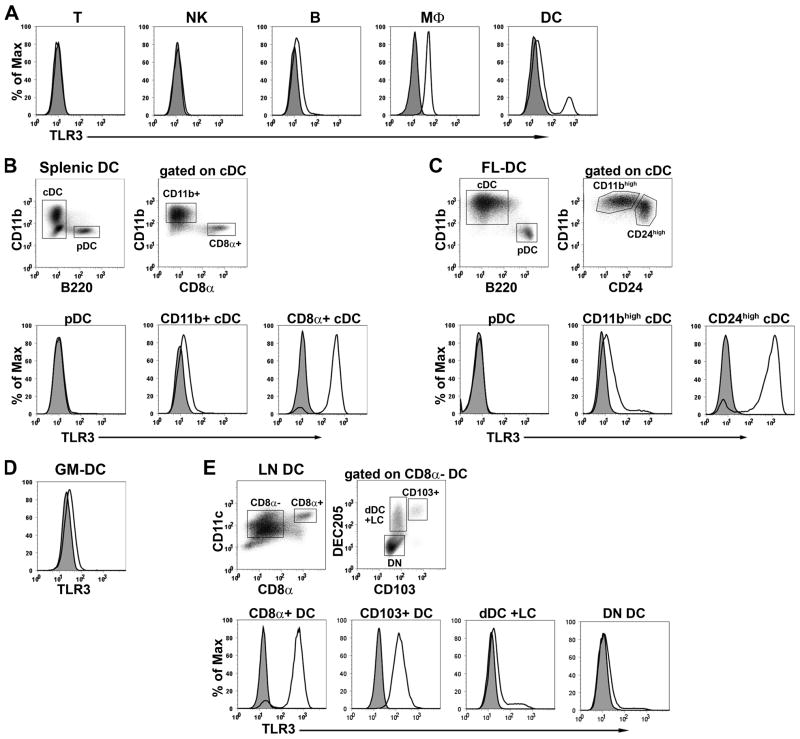
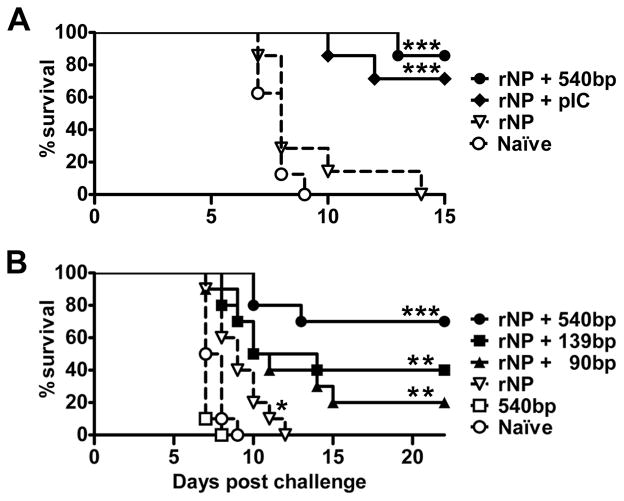
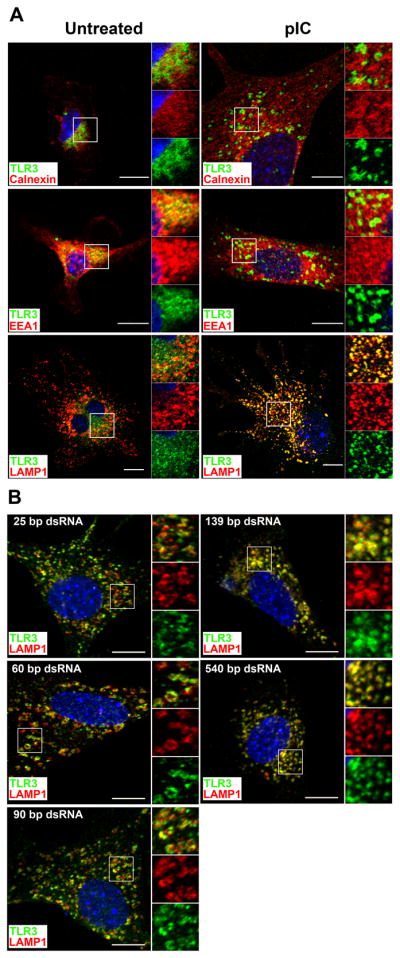
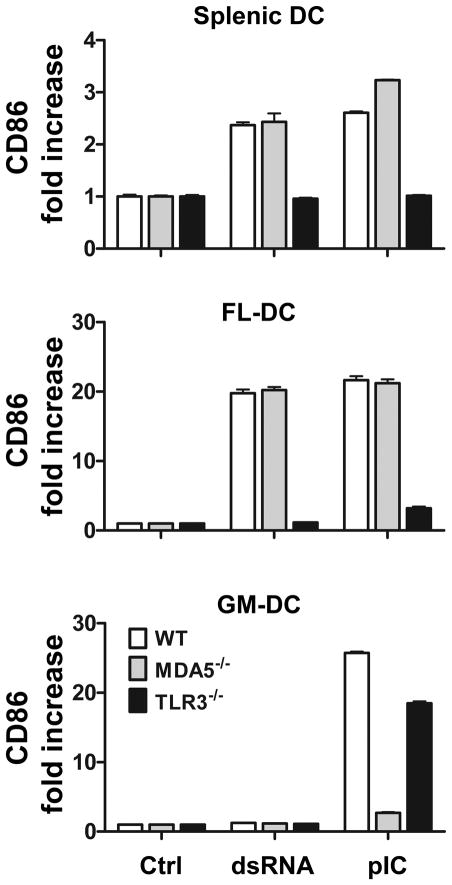
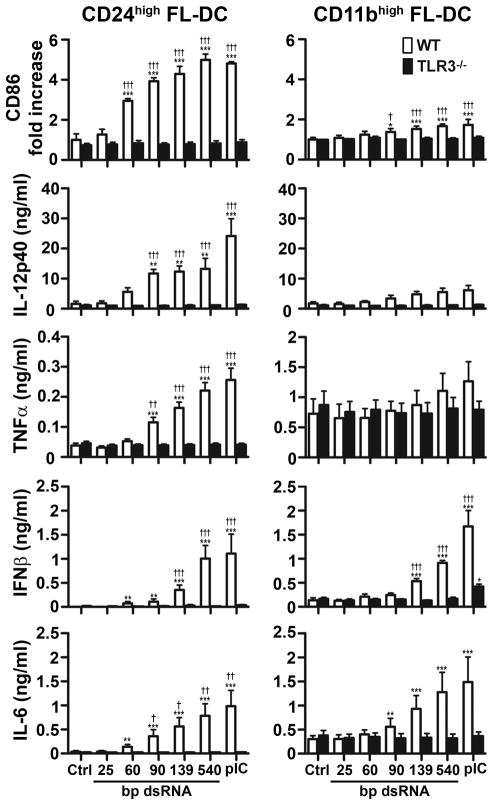
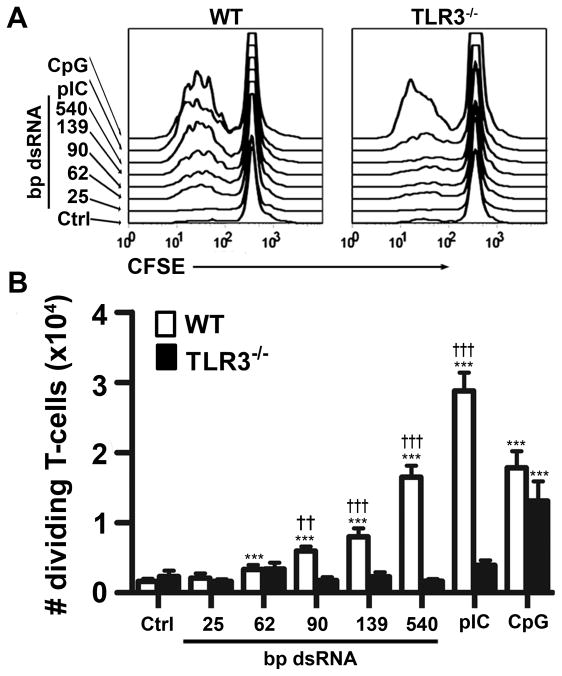
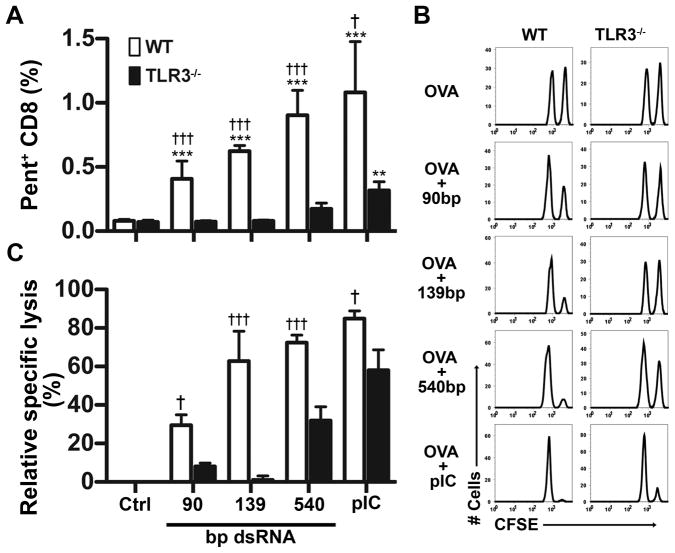
Maturation of dendritic cells (DC) to competent APC is essential for the generation of acquired immunity and is a major function of adjuvants. dsRNA, a molecular signature of viral infection, drives DC maturation by activating TLR3, but the size of dsRNA required to activate DC and the expression patterns of TLR3 protein in DC subsets have not been established. In this article, we show that cross-priming CD8α(+) and CD103(+) DC subsets express much greater levels of TLR3 than other DC. In resting DC, TLR3 is located in early endosomes and other intracellular compartments but migrates to LAMP1(+) endosomes on stimulation with a TLR3 ligand. Using homogeneous dsRNA oligonucleotides (ONs) ranging in length from 25 to 540 bp, we observed that a minimum length of ∼90 bp was sufficient to induce CD86, IL-12p40, IFN-β, TNF-α, and IL-6 expression, and to mature DC into APC that cross-presented exogenous Ags to CD8(+) T cells. TLR3 was essential for activation of DC by dsRNA ONs, and the potency of activation increased with dsRNA length and varied between DC subsets. In vivo, dsRNA ONs, in a size-dependent manner, served as adjuvants for the generation of Ag-specific CTL and for inducing protection against lethal challenge with influenza virus when given with influenza nucleoprotein as an immunogen. These results provide the basis for the development of TLR3-specific adjuvants capable of inducing immune responses tailored for viral pathogens.
树突状细胞(DC)成熟为有功能的抗原呈递细胞(APC)对于获得性免疫的产生至关重要,并且是佐剂的主要功能。双链RNA(dsRNA)作为病毒感染的分子标志,通过激活Toll样受体3(TLR3)来驱动DC成熟,但激活DC所需的dsRNA大小以及DC亚群中TLR3蛋白的表达模式尚未明确。在本文中,我们发现交叉呈递的CD8α(+)和CD103(+) DC亚群表达的TLR3水平远高于其他DC。在静息DC中,TLR3位于早期内体和其他细胞内区室,但在用TLR3配体刺激后会迁移至溶酶体相关膜蛋白1(LAMP1)(+)内体。使用长度范围为25至540 bp的均一化dsRNA寡核苷酸(ONs),我们观察到约90 bp的最小长度足以诱导CD86、白细胞介素12p40(IL-12p40)、干扰素-β(IFN-β)、肿瘤坏死因子-α(TNF-α)和白细胞介素-6(IL-6)的表达,并使DC成熟为能够将外源性抗原交叉呈递给CD8(+) T细胞的APC。TLR3对于dsRNA ONs激活DC至关重要,激活效力随dsRNA长度增加而增强,且在DC亚群之间存在差异。在体内,dsRNA ONs以大小依赖的方式作为佐剂,与流感核蛋白作为免疫原一起使用时,可用于产生抗原特异性细胞毒性T淋巴细胞(CTL)并诱导对流感病毒致死性攻击的保护作用。这些结果为开发能够诱导针对病毒病原体的定制免疫反应的TLR3特异性佐剂提供了依据。