Department of Genetics, University of Pennsylvania School of Medicine, Philadelphia, PA 19104, USA.
Neuroscience. 2011 May 5;181:48-66. doi: 10.1016/j.neuroscience.2011.02.029. Epub 2011 Feb 17.
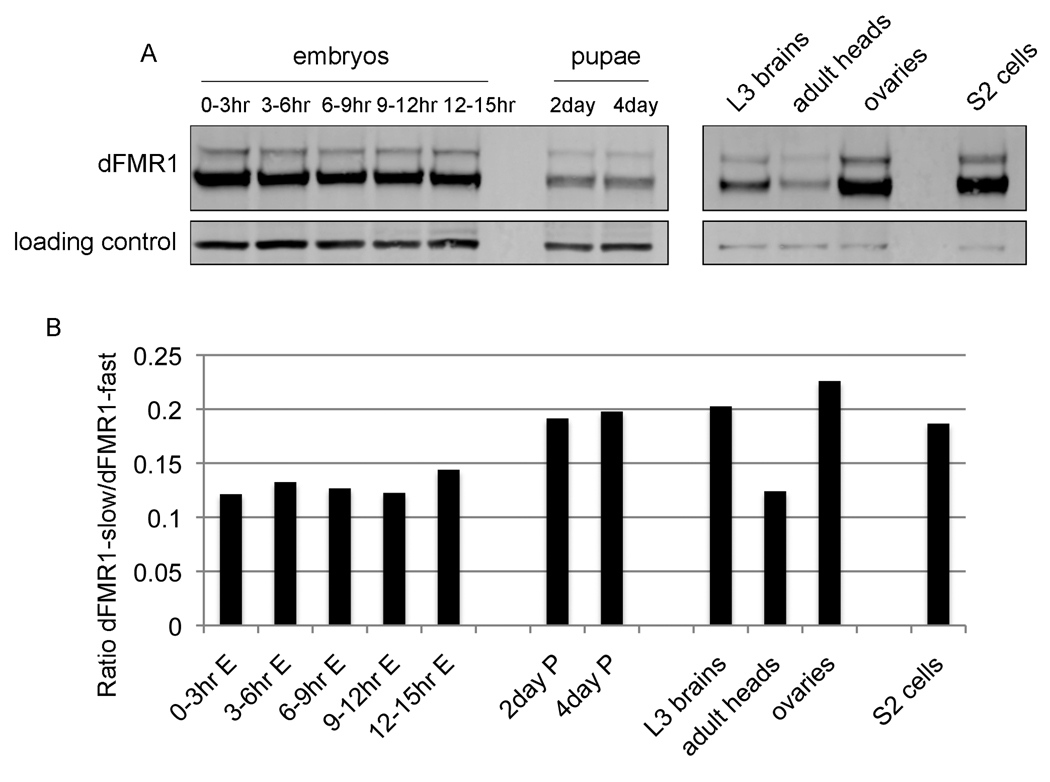
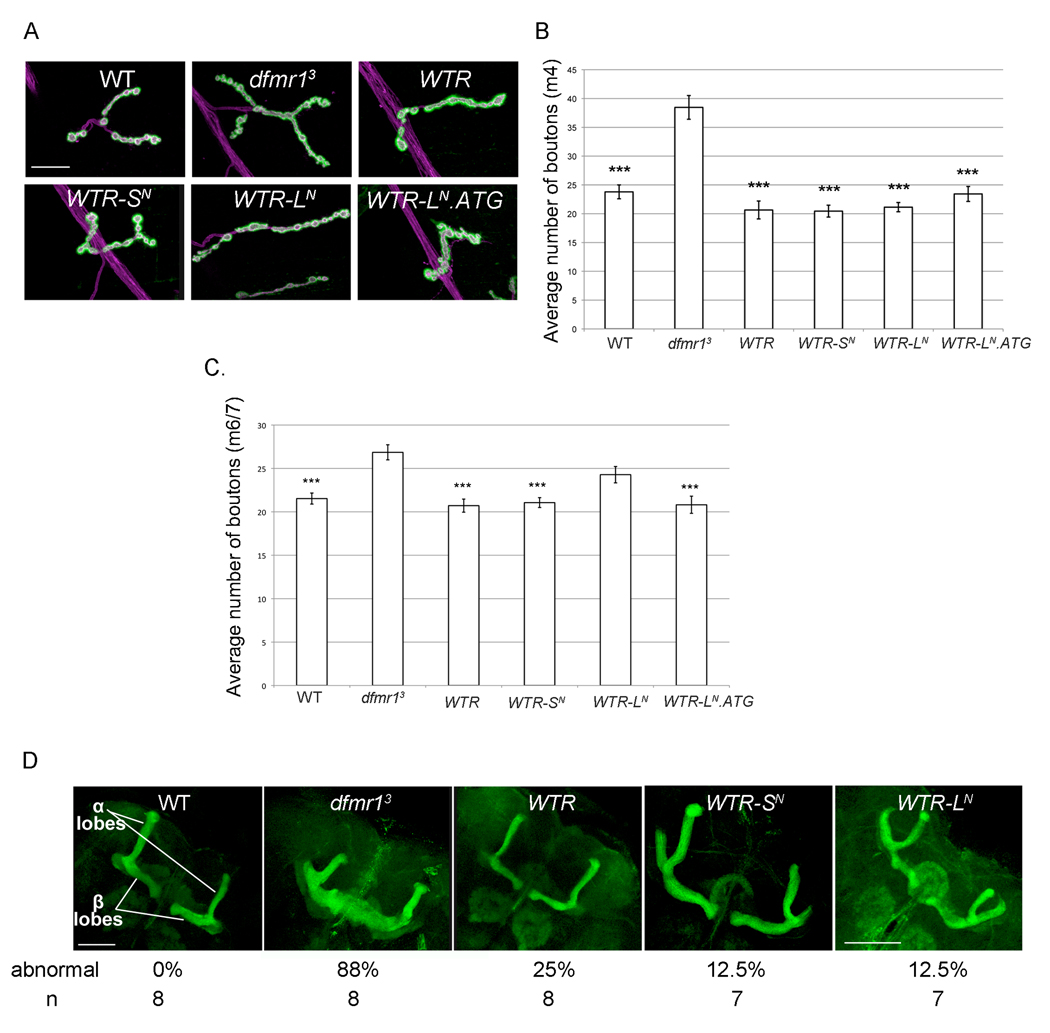
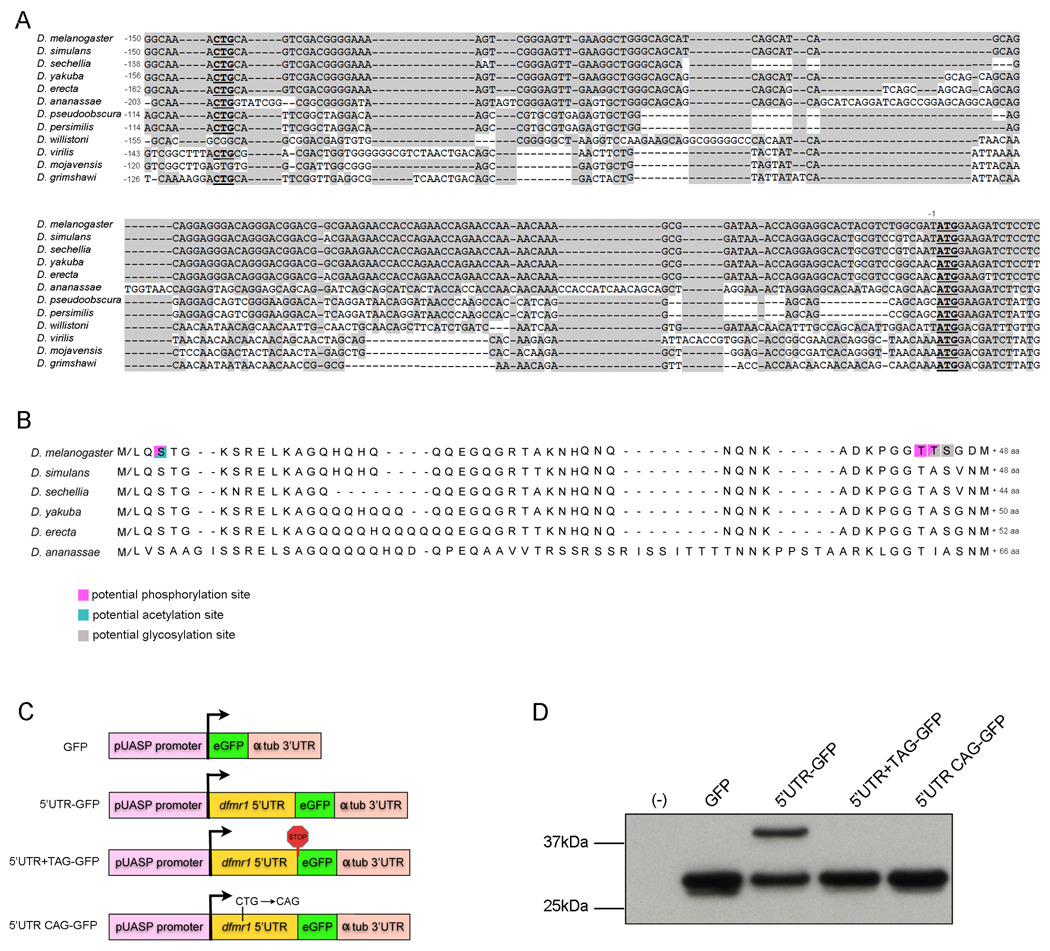
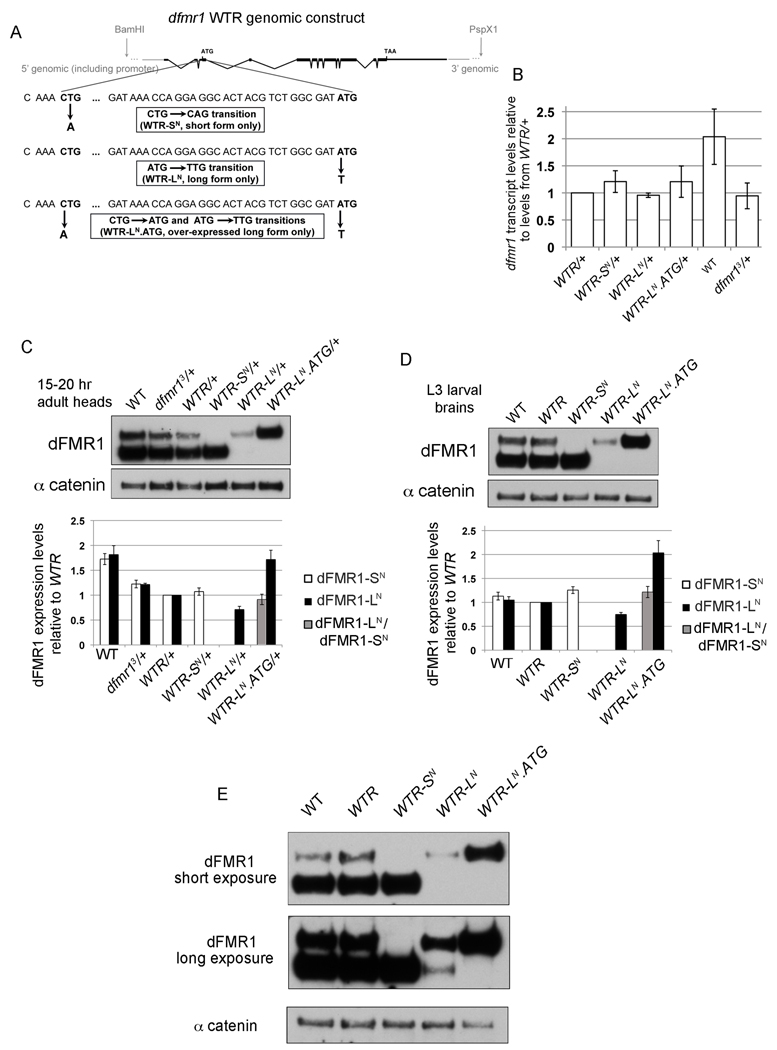
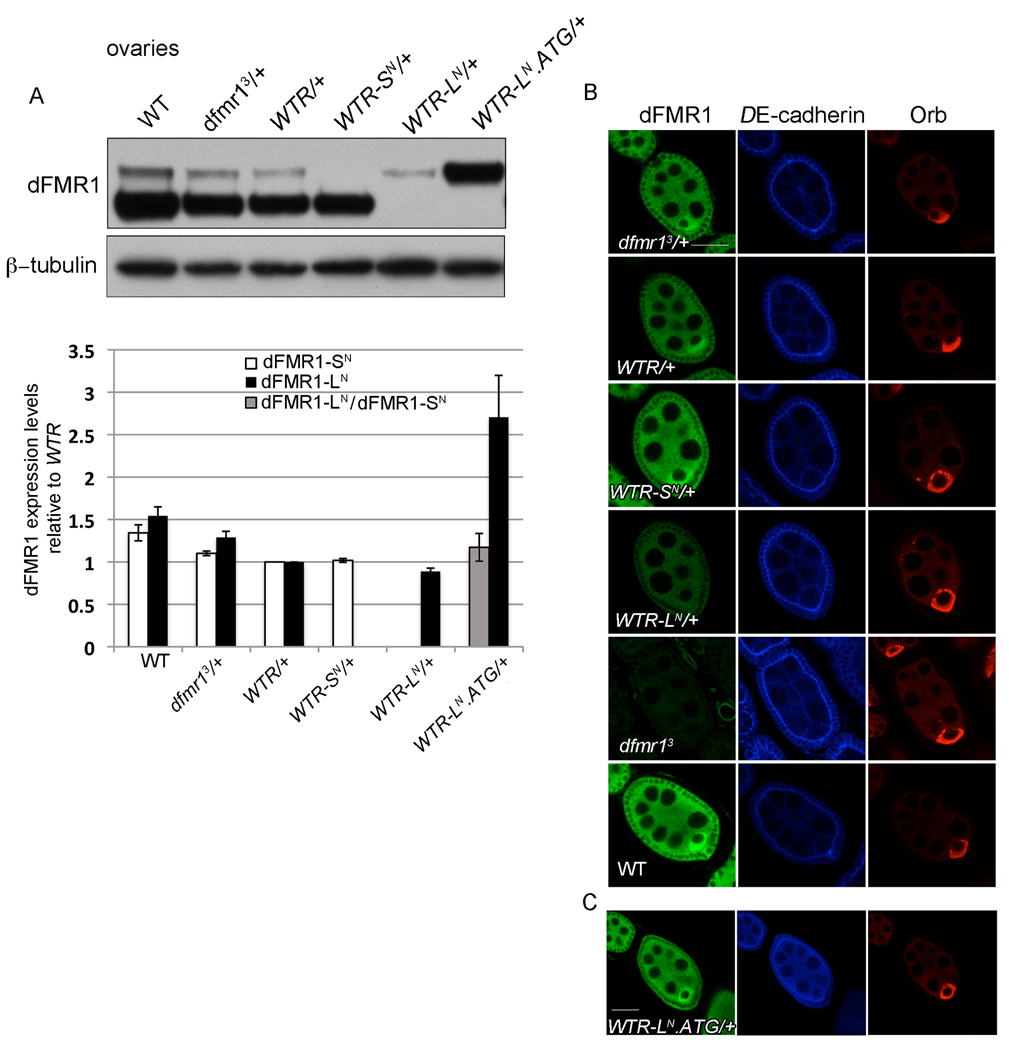
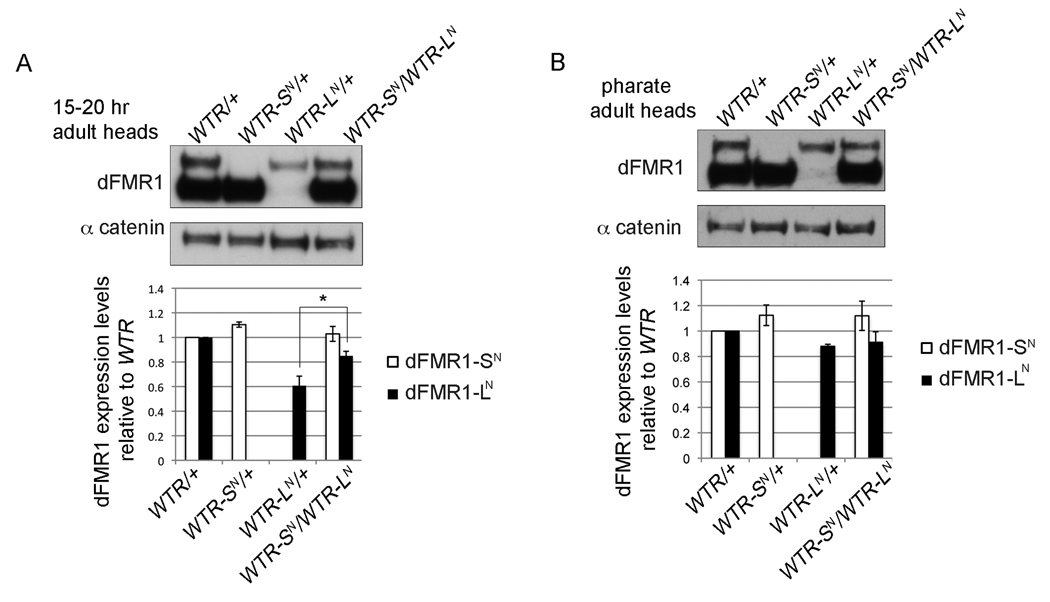
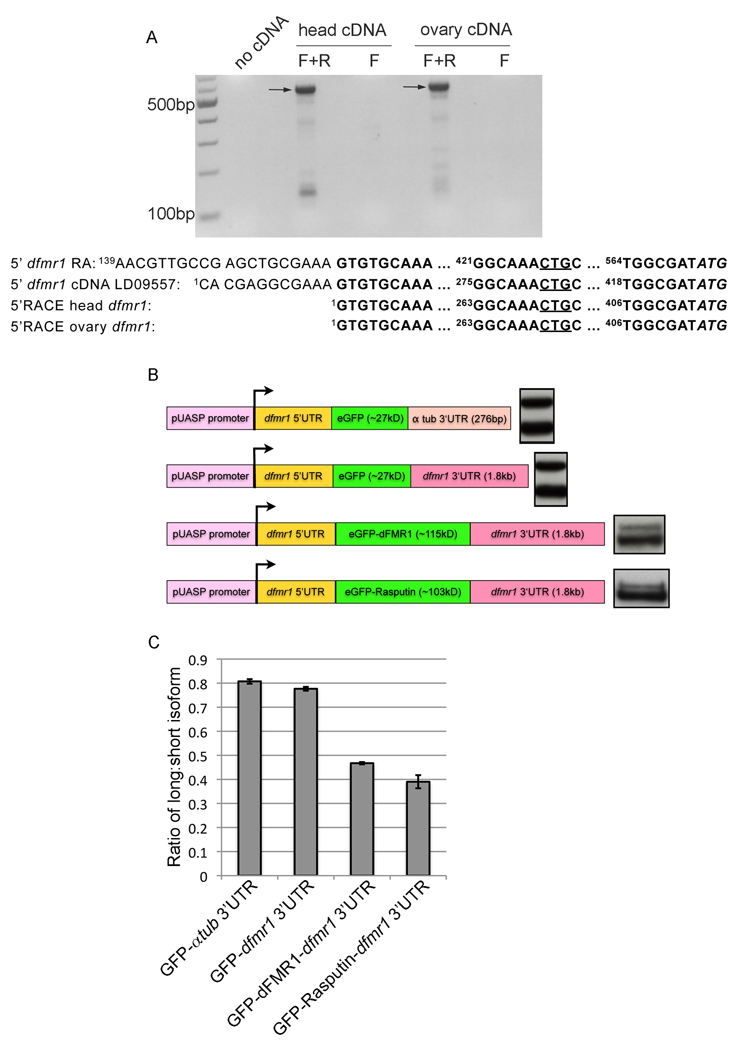
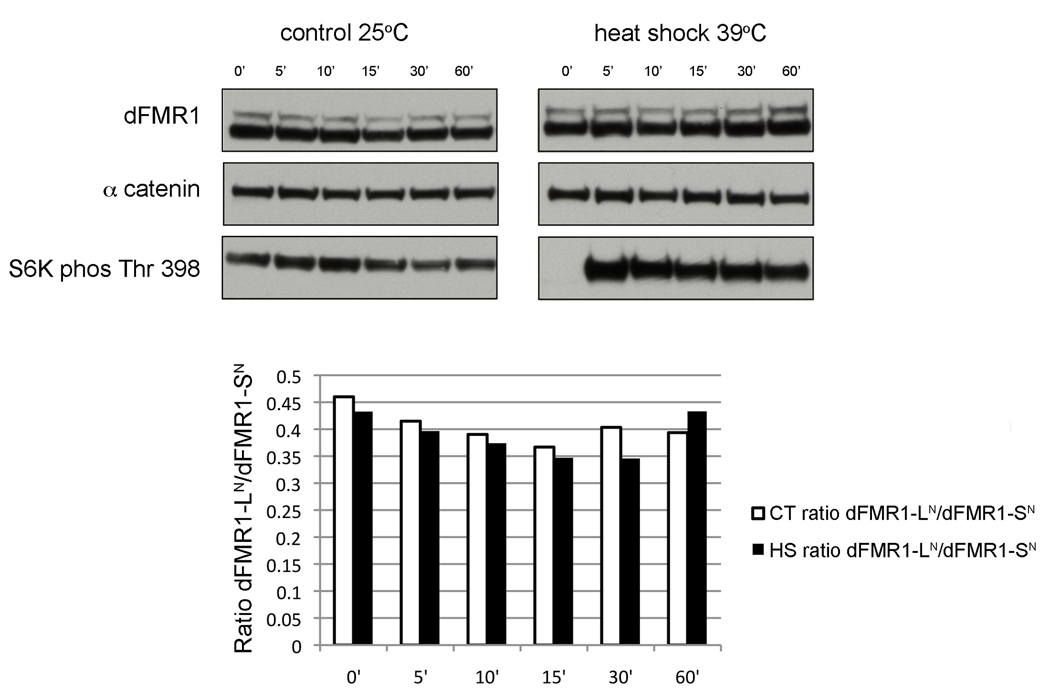
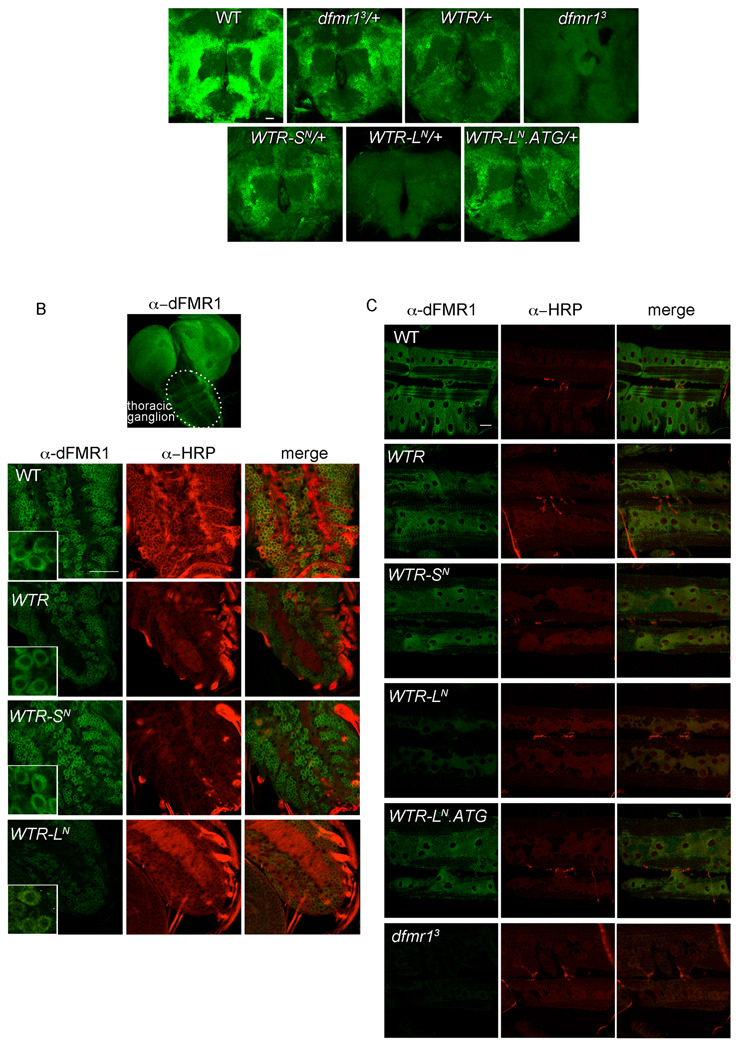
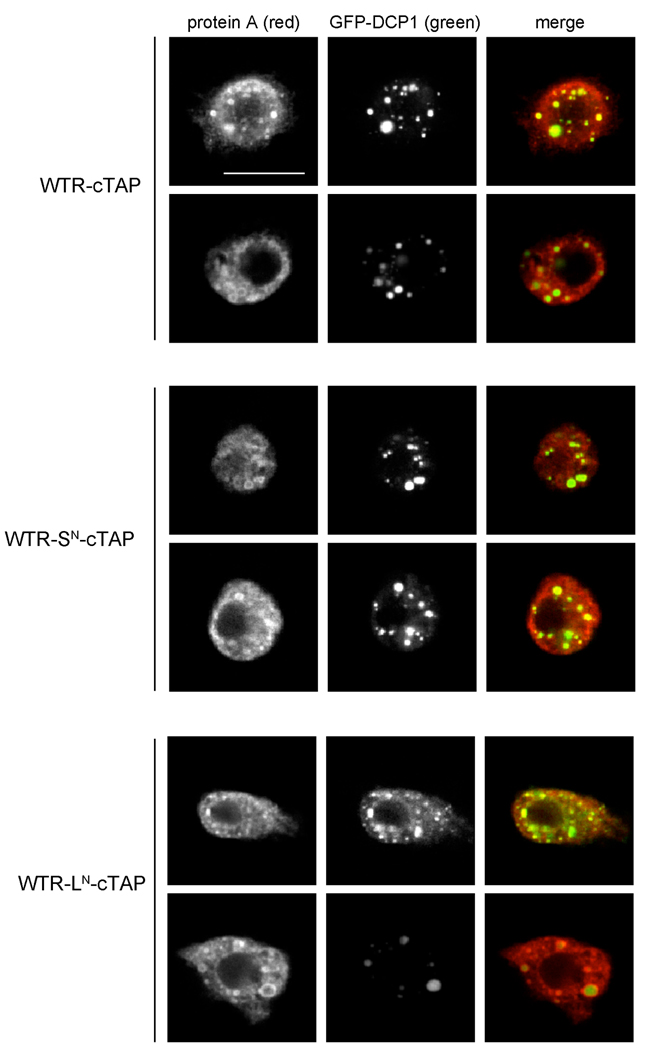
Fragile X syndrome is caused by the loss of expression of the fragile X mental retardation protein (FMRP). As a RNA binding protein, FMRP functions in translational regulation, localization, and stability of its neuronal target transcripts. The Drosophila homologue, dFMR1, is well conserved in sequence and function with respect to human FMRP. Although dFMR1 is known to express two main isoforms, the mechanism behind production of the second, more slowly migrating isoform has remained elusive. Furthermore, it remains unknown whether the two isoforms may also contribute differentially to dFMR1 function. We have found that this second dFMR1 isoform is generated through an alternative translational start site in the dfmr1 5'UTR. This 5'UTR coding sequence is well conserved in the melanogaster group. Translation of the predominant, smaller form of dFMR1 (dFMR1-S(N)) begins at a canonical start codon (ATG), whereas translation of the minor, larger form (dFMR1-L(N)) begins upstream at a non-canonical start codon (CTG). To assess the contribution of the N-terminal extension toward dFMR1 activity, we generated transgenic flies that exclusively express either dFMR1-S(N) or dFMR1-L(N). Expression analyses throughout development revealed that dFMR1-S(N) is required for normal dFMR1-L(N) expression levels in adult brains. In situ expression analyses showed that either dFMR1-S(N) or dFMR1-L(N) is individually sufficient for proper dFMR1 localization in the nervous system. Functional studies demonstrated that both dFMR1-S(N) and dFMR1-L(N) can function independently to rescue dfmr1 null defects in synaptogenesis and axon guidance. Thus, dfmr1 encodes two functional isoforms with respect to expression and activity throughout neuronal development.
脆性 X 综合征是由脆性 X 智力低下蛋白(FMRP)表达缺失引起的。作为一种 RNA 结合蛋白,FMRP 参与其神经元靶标转录本的翻译调控、定位和稳定性。果蝇同源物 dFMR1 在序列和功能上与人 FMRP 高度保守。尽管已知 dFMR1 表达两种主要的同工型,但产生第二种迁移速度较慢的同工型的机制仍不清楚。此外,两种同工型是否也可能对 dFMR1 功能产生不同的影响也不得而知。我们发现这种第二种 dFMR1 同工型是通过 dfmr1 5'UTR 中的一个替代翻译起始位点产生的。这个 5'UTR 编码序列在黑腹果蝇组中高度保守。主要的较小形式 dFMR1(dFMR1-S(N))的翻译从一个典型的起始密码子(ATG)开始,而较小的较大形式(dFMR1-L(N))的翻译从上游的非典型起始密码子(CTG)开始。为了评估 N 端延伸对 dFMR1 活性的贡献,我们生成了专门表达 dFMR1-S(N)或 dFMR1-L(N)的转基因果蝇。在整个发育过程中的表达分析表明,dFMR1-S(N)是成年大脑中正常表达 dFMR1-L(N)所必需的。原位表达分析表明,dFMR1-S(N)或 dFMR1-L(N)中的任何一种都足以使 dFMR1 在神经系统中的定位正常。功能研究表明,dFMR1-S(N)和 dFMR1-L(N)都可以独立发挥作用,以挽救 dfmr1 缺失在突触发生和轴突导向中的缺陷。因此,dfmr1 编码两种具有表达和活性的功能同工型,贯穿神经元发育的始终。