Department of Physics, University of South Florida, Tampa, Florida, United States of America.
PLoS One. 2011 Apr 5;6(4):e18171. doi: 10.1371/journal.pone.0018171.
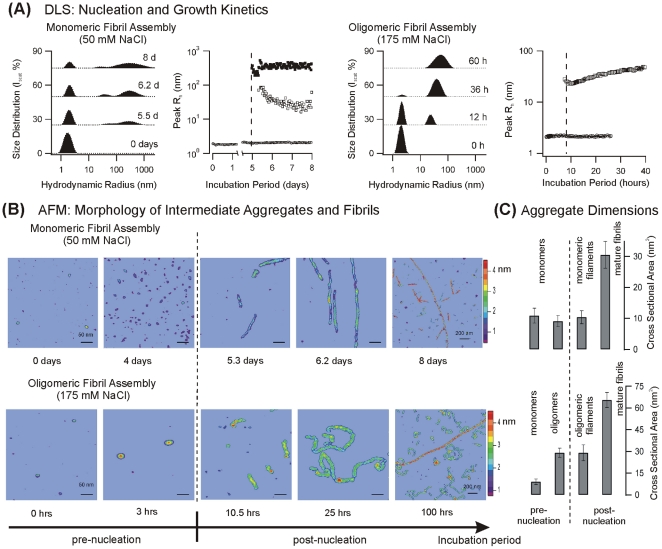
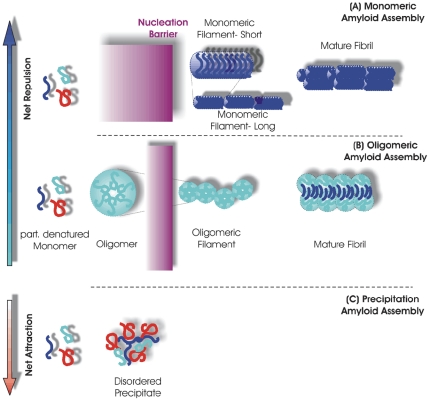
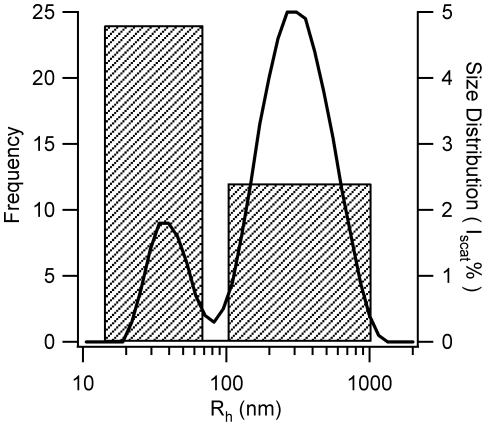
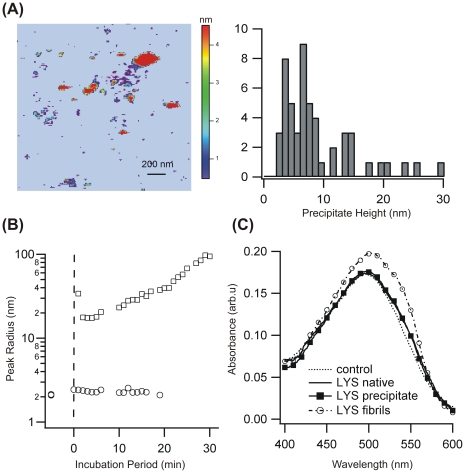
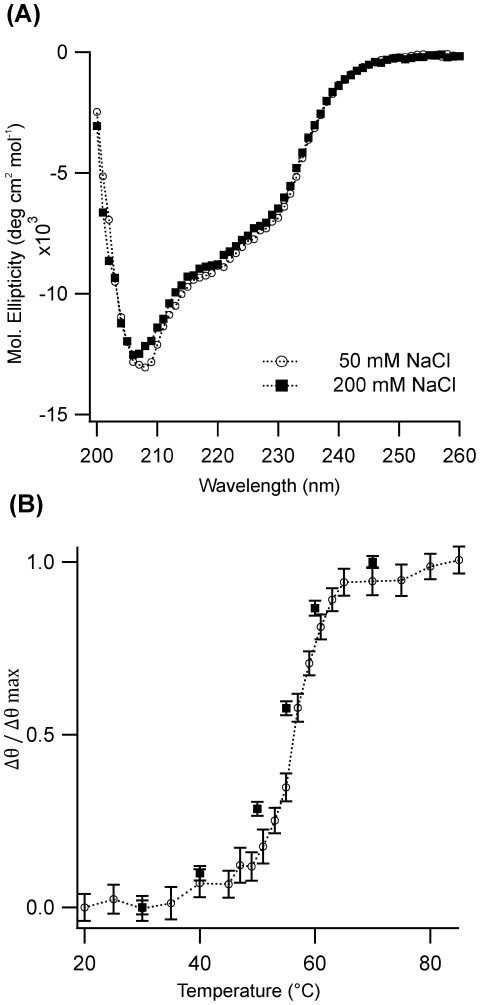
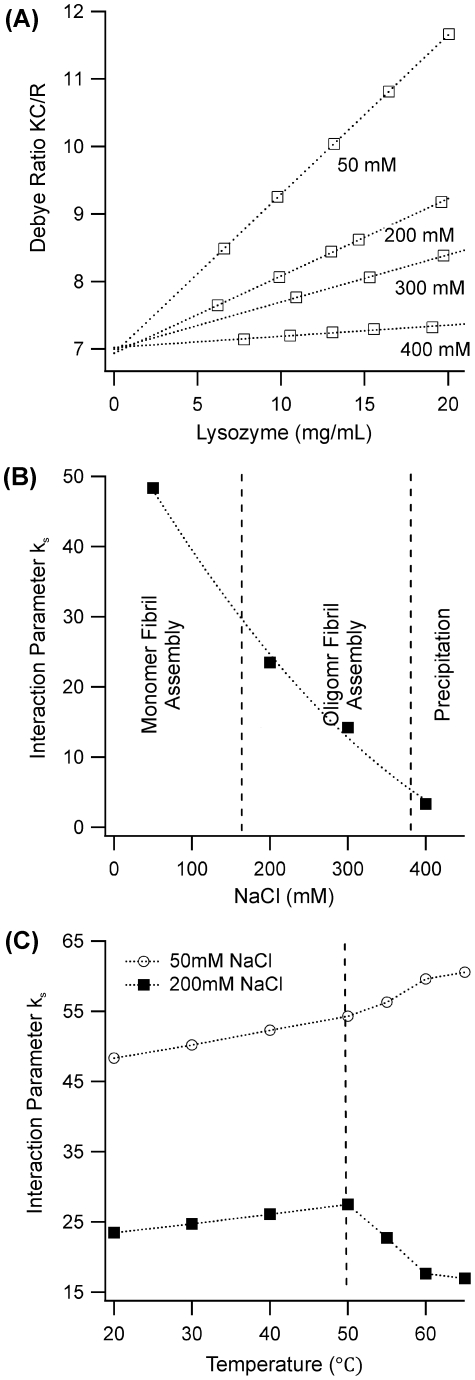
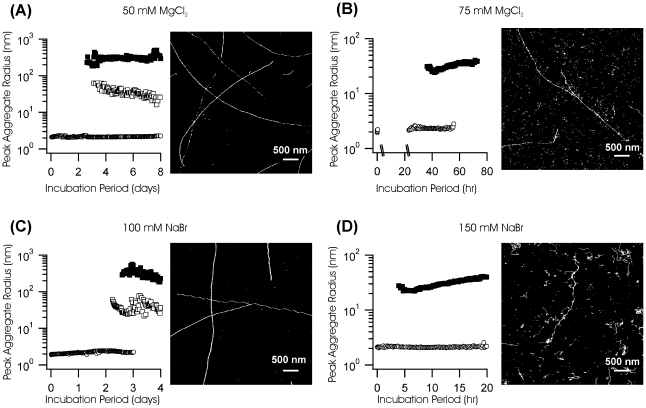
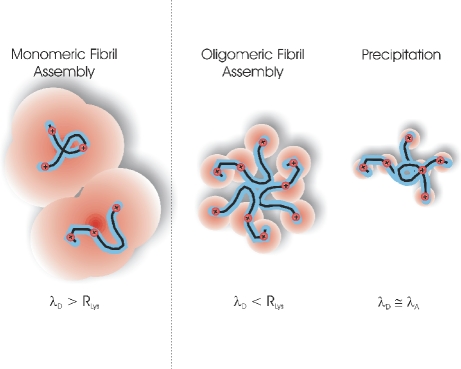
Formation of large protein fibrils with a characteristic cross β-sheet architecture is the key indicator for a wide variety of systemic and neurodegenerative amyloid diseases. Recent experiments have strongly implicated oligomeric intermediates, transiently formed during fibril assembly, as critical contributors to cellular toxicity in amyloid diseases. At the same time, amyloid fibril assembly can proceed along different assembly pathways that might or might not involve such oligomeric intermediates. Elucidating the mechanisms that determine whether fibril formation proceeds along non-oligomeric or oligomeric pathways, therefore, is important not just for understanding amyloid fibril assembly at the molecular level but also for developing new targets for intervening with fibril formation. We have investigated fibril formation by hen egg white lysozyme, an enzyme for which human variants underlie non-neuropathic amyloidosis. Using a combination of static and dynamic light scattering, atomic force microscopy and circular dichroism, we find that amyloidogenic lysozyme monomers switch between three different assembly pathways: from monomeric to oligomeric fibril assembly and, eventually, disordered precipitation as the ionic strength of the solution increases. Fibril assembly only occurred under conditions of net repulsion among the amyloidogenic monomers while net attraction caused precipitation. The transition from monomeric to oligomeric fibril assembly, in turn, occurred as salt-mediated charge screening reduced repulsion among individual charged residues on the same monomer. We suggest a model of amyloid fibril formation in which repulsive charge interactions are a prerequisite for ordered fibril assembly. Furthermore, the spatial extent of non-specific charge screening selects between monomeric and oligomeric assembly pathways by affecting which subset of denatured states can form suitable intermolecular bonds and by altering the energetic and entropic requirements for the initial intermediates emerging along the monomeric vs. oligomeric assembly path.
形成具有特征性交叉β-折叠结构的大型蛋白质纤维是各种系统性和神经退行性淀粉样疾病的关键指标。最近的实验强烈表明,寡聚中间体在纤维组装过程中短暂形成,是淀粉样疾病细胞毒性的关键贡献者。与此同时,淀粉样纤维的组装可以沿着不同的组装途径进行,这些途径可能涉及也可能不涉及这种寡聚中间体。因此,阐明决定纤维形成是沿着非寡聚途径还是寡聚途径进行的机制,不仅对于理解淀粉样纤维在分子水平上的组装很重要,而且对于开发干预纤维形成的新靶点也很重要。我们研究了鸡卵清溶菌酶的纤维形成,这种酶的人类变体是神经病变淀粉样变性的基础。我们使用静态和动态光散射、原子力显微镜和圆二色性的组合,发现淀粉样溶菌酶单体在三种不同的组装途径之间切换:从单体到寡聚纤维组装,最终在溶液离子强度增加时无序沉淀。只有在淀粉样单体之间存在净排斥的情况下,纤维组装才会发生,而净吸引则会导致沉淀。单体到寡聚纤维组装的转变反过来又发生在盐介导的电荷屏蔽减少了同一单体上带电残基之间的排斥作用时。我们提出了一种淀粉样纤维形成的模型,其中排斥电荷相互作用是有序纤维组装的前提。此外,非特异性电荷屏蔽的空间范围通过影响哪些变性状态可以形成合适的分子间键,以及通过改变沿单体与寡聚组装路径出现的初始中间物的能量和熵要求,在单体与寡聚组装途径之间进行选择。