Department of Biochemistry and Molecular Biology, University of Texas Medical Branch at Galveston, Galveston, Texas, USA.
mBio. 2011 May 17;2(3):e00079-11. doi: 10.1128/mBio.00079-11. Print 2011.
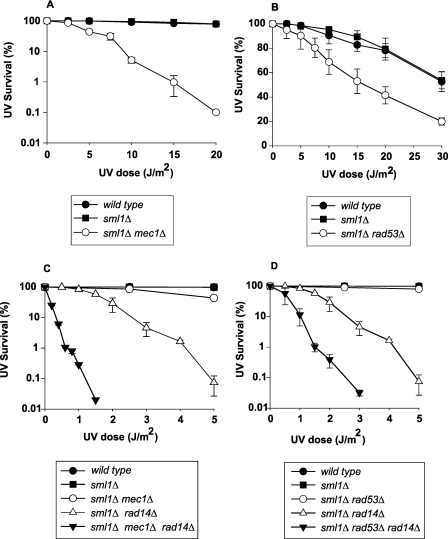
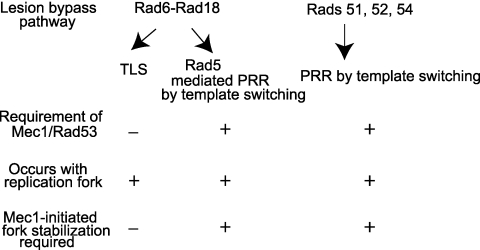
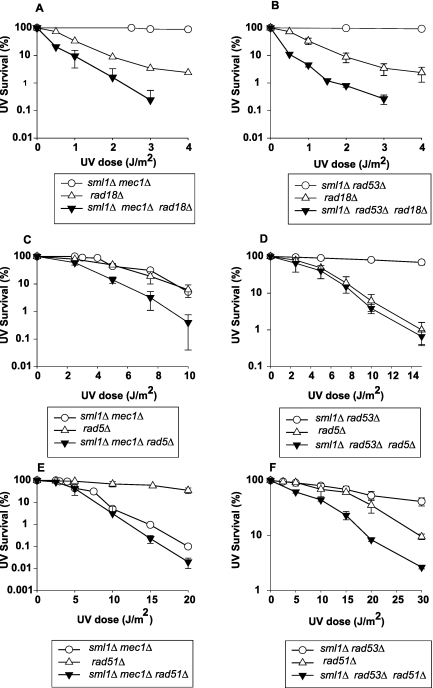
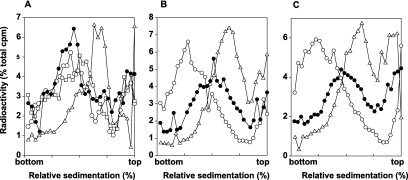
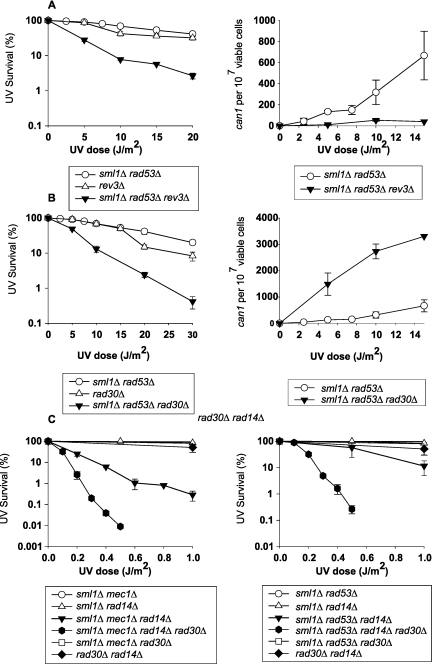
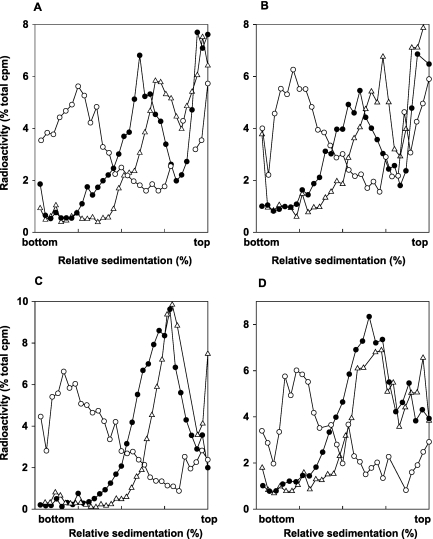
DNA lesions in the template strand block the replication fork. In Saccharomyces cerevisiae, replication through DNA lesions occurs via a Rad6/Rad18-dependent pathway where lesions can be bypassed by the action of translesion synthesis (TLS) DNA polymerases η and ζ or by Rad5-mediated template switching. An alternative Rad6/Rad18-independent but Rad52-dependent template switching pathway can also restore the continuity of the replication fork. The Mec1/Rad53-dependent replication checkpoint plays a crucial role in the maintenance of stable and functional replication forks in yeast cells with DNA damage; however, it has remained unclear which of the lesion bypass processes requires the activation of replication checkpoint-mediated fork stabilization. Here we show that postreplication repair (PRR) of newly synthesized DNA in UV-damaged yeast cells is inhibited in the absence of Mec1 and Rad53 proteins. Since TLS remains functional in cells lacking these checkpoint kinases and since template switching by the Rad5 and Rad52 pathways provides the alternative means of lesion bypass and requires Mec1/Rad53, we infer that lesion bypass by the template switching pathways occurs in conjunction with the replication fork that has been stabilized at the lesion site by the action of Mec1/Rad53-mediated replication checkpoint.
Eukaryotic cells possess mechanisms called checkpoints that act to stop the cell cycle when DNA replication is halted by lesions in the template strand. Upon stalling of the ongoing replication at the lesion site, the recruitment of Mec1 and Rad53 kinases to the replication ensemble initiates the checkpoint wherein Mec1-mediated phosphorylation of Rad53 activates the pathway. A crucial role of replication checkpoint is to stabilize the replication fork by maintaining the association of DNA polymerases with the other replication components at the stall site. Our observations that Mec1 and Rad53 are required for lesion bypass by template switching have important implications for whether lesion bypass occurs in conjunction with the stalled replication ensemble or in gaps that could have been left behind the newly restarted forks. We discuss this important issue and suggest that lesion bypass in Saccharomyces cerevisiae cells occurs in conjunction with the stalled replication forks and not in gaps.
模板链上的 DNA 损伤会阻碍复制叉。在酿酒酵母中,通过 Rad6/Rad18 依赖性途径进行 DNA 损伤部位的复制,在此过程中,损伤部位可以通过跨损伤合成(TLS)DNA 聚合酶 η 和 ζ 的作用或 Rad5 介导的模板转换来绕过。另一种 Rad6/Rad18 非依赖性但 Rad52 依赖性的模板转换途径也可以恢复复制叉的连续性。Mec1/Rad53 依赖性复制检查点在维持酵母细胞中具有 DNA 损伤的稳定和功能性复制叉方面起着至关重要的作用;然而,尚不清楚哪种损伤绕过过程需要激活复制检查点介导的叉稳定。在这里,我们表明,在缺乏 Mec1 和 Rad53 蛋白的情况下,紫外线损伤酵母细胞中新合成 DNA 的复制后修复(PRR)被抑制。由于在缺乏这些检查点激酶的细胞中 TLS 仍然具有功能,并且 Rad5 和 Rad52 途径的模板转换提供了损伤绕过的替代方法,并且需要 Mec1/Rad53,我们推断损伤绕过通过模板转换途径发生在由 Mec1/Rad53 介导的复制检查点作用于损伤部位的复制叉。
真核细胞具有称为检查点的机制,当模板链上的损伤导致 DNA 复制停滞时,这些机制会使细胞周期停止。在损伤部位的正在进行的复制停滞时,Mec1 和 Rad53 激酶募集到复制复合物中启动检查点,其中 Mec1 介导的 Rad53 磷酸化激活途径。复制检查点的一个关键作用是通过维持 DNA 聚合酶与停滞部位其他复制成分的结合来稳定复制叉。我们的观察结果表明,Mec1 和 Rad53 对于通过模板转换进行损伤绕过是必需的,这对于损伤绕过是发生在停滞的复制复合物中还是在新重新启动的叉后面可能留下的间隙中具有重要意义。我们讨论了这个重要问题,并提出在酿酒酵母细胞中,损伤绕过发生在停滞的复制叉上,而不是在间隙中。