Department of Microbiology and Molecular Genetics, Harvard Medical School, Boston, Massachusetts, United States of America.
PLoS Pathog. 2011 Jun;7(6):e1002073. doi: 10.1371/journal.ppat.1002073. Epub 2011 Jun 2.
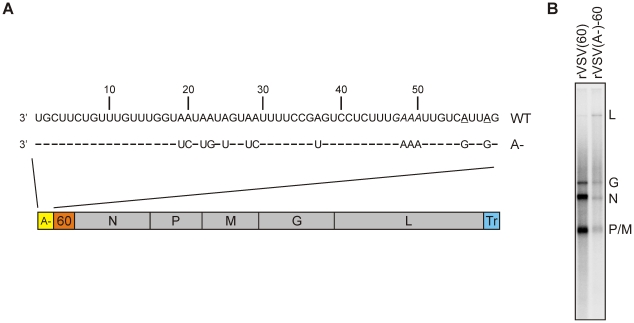
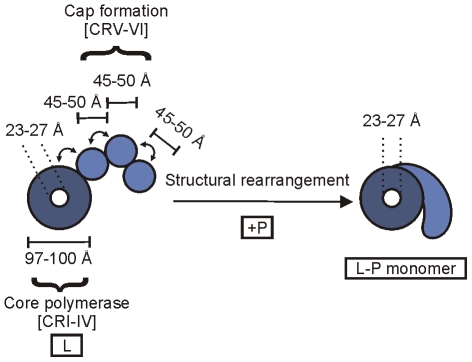
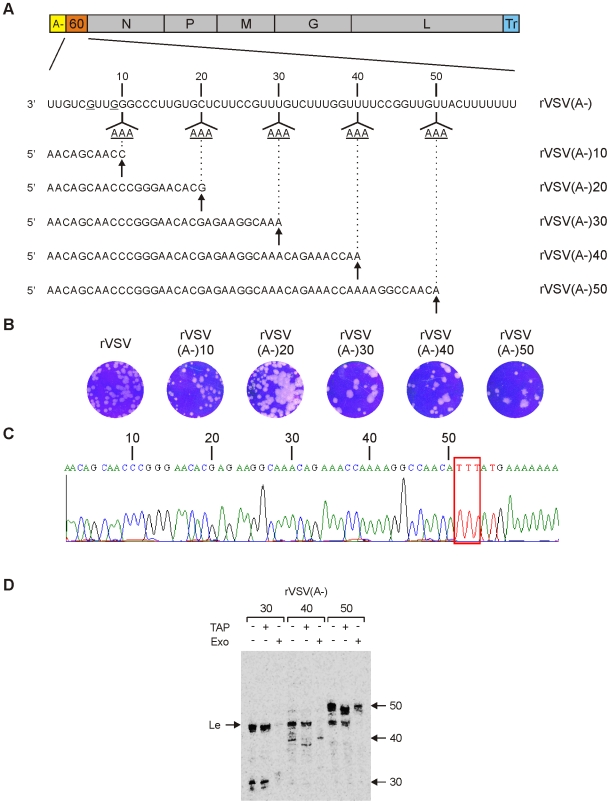
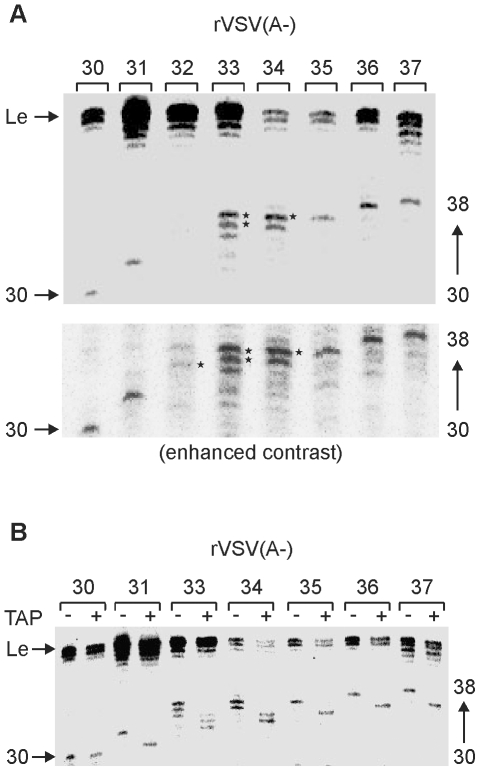
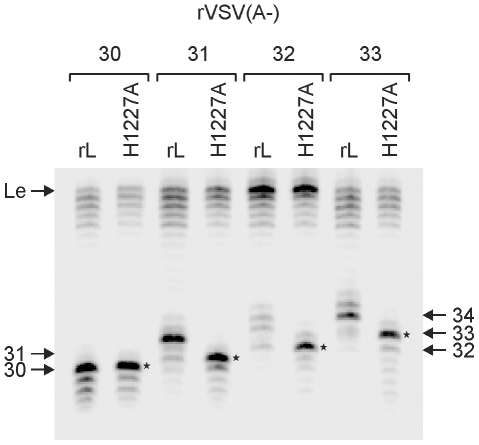
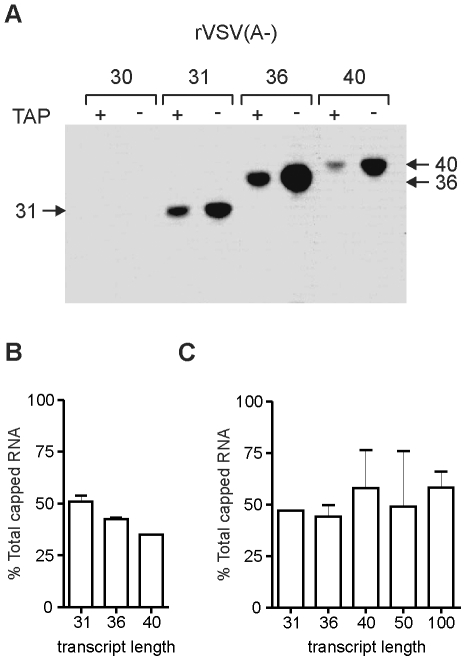
The RNA synthesis machinery of vesicular stomatitis virus (VSV) comprises the genomic RNA encapsidated by the viral nucleocapsid protein (N) and associated with the RNA dependent RNA polymerase, the viral components of which are a large protein (L) and an accessory phosphoprotein (P). The 241 kDa L protein contains all the enzymatic activities necessary for synthesis of the viral mRNAs, including capping, cap methylation and polyadenylation. Those RNA processing reactions are intimately coordinated with nucleotide polymerization such that failure to cap results in termination of transcription and failure to methylate can result in hyper polyadenylation. The mRNA processing reactions thus serve as a critical check point in viral RNA synthesis which may control the synthesis of incorrectly modified RNAs. Here, we report the length at which viral transcripts first gain access to the capping machinery during synthesis. By reconstitution of transcription in vitro with highly purified recombinant polymerase and engineered templates in which we omitted sites for incorporation of UTP, we found that transcripts that were 30-nucleotides in length were uncapped, whereas those that were 31-nucleotides in length contained a cap structure. The minimal RNA length required for mRNA cap addition was also sufficient for methylation since the 31-nucleotide long transcripts were methylated at both ribose-2'-O and guanine-N-7 positions. This work provides insights into the spatial relationship between the active sites for the RNA dependent RNA polymerase and polyribonucleotidyltransferase responsible for capping of the viral RNA. We combine the present findings with our recently described electron microscopic structure of the VSV polymerase and propose a model of how the spatial arrangement of the capping activities of L may influence nucleotide polymerization.
水疱性口炎病毒 (VSV) 的 RNA 合成机制包括基因组 RNA 被病毒核衣壳蛋白 (N) 包裹,并与 RNA 依赖性 RNA 聚合酶相关联,该聚合酶的病毒成分是一种大蛋白 (L) 和一种辅助磷蛋白 (P)。241kDa 的 L 蛋白包含合成病毒 mRNAs 所需的所有酶活性,包括加帽、帽甲基化和多聚腺苷酸化。这些 RNA 加工反应与核苷酸聚合密切协调,以至于加帽失败会导致转录终止,而甲基化失败可能导致过度多聚腺苷酸化。因此,mRNA 加工反应是病毒 RNA 合成中的一个关键检查点,它可能控制了不正确修饰的 RNA 的合成。在这里,我们报告了病毒转录物在合成过程中首次获得加帽机制的长度。通过在高度纯化的重组聚合酶和我们省略 UTP 掺入位点的工程模板中体外重建转录,我们发现长度为 30 个核苷酸的转录物未加帽,而长度为 31 个核苷酸的转录物含有帽结构。用于 mRNA 帽添加的最小 RNA 长度也足以进行甲基化,因为 31 个核苷酸长的转录物在核糖-2'-O 和鸟嘌呤-N-7 位置都被甲基化。这项工作深入了解了负责病毒 RNA 加帽的 RNA 依赖性 RNA 聚合酶和多核糖核苷酸转移酶的活性位点之间的空间关系。我们将目前的发现与我们最近描述的 VSV 聚合酶的电子显微镜结构结合起来,提出了 L 的加帽活性的空间排列如何影响核苷酸聚合的模型。