Department of Biochemistry and Molecular Biology, Colorado State University, Fort Collins, Colorado, United States of America.
PLoS One. 2011;6(7):e21953. doi: 10.1371/journal.pone.0021953. Epub 2011 Jul 8.
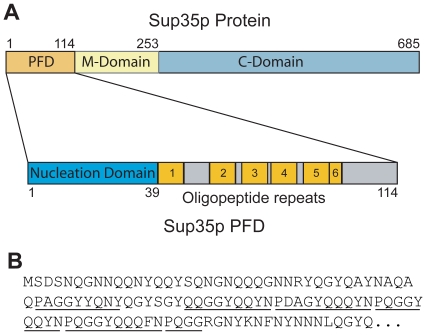
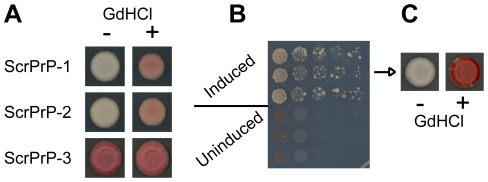
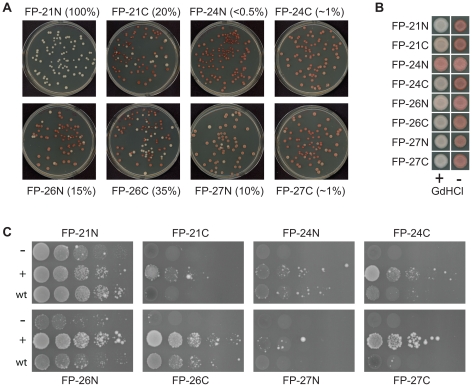
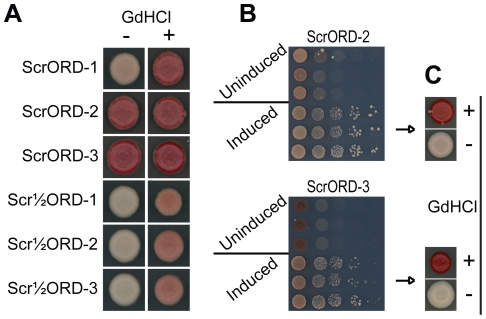
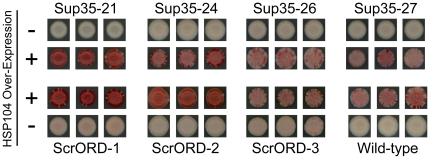
[PSI(+)], the prion form of the yeast Sup35 protein, results from the structural conversion of Sup35 from a soluble form into an infectious amyloid form. The infectivity of prions is thought to result from chaperone-dependent fiber cleavage that breaks large prion fibers into smaller, inheritable propagons. Like the mammalian prion protein PrP, Sup35 contains an oligopeptide repeat domain. Deletion analysis indicates that the oligopeptide repeat domain is critical for [PSI(+)] propagation, while a distinct region of the prion domain is responsible for prion nucleation. The PrP oligopeptide repeat domain can substitute for the Sup35 oligopeptide repeat domain in supporting [PSI(+)] propagation, suggesting a common role for repeats in supporting prion maintenance. However, randomizing the order of the amino acids in the Sup35 prion domain does not block prion formation or propagation, suggesting that amino acid composition is the primary determinant of Sup35's prion propensity. Thus, it is unclear what role the oligopeptide repeats play in [PSI(+)] propagation: the repeats could simply act as a non-specific spacer separating the prion nucleation domain from the rest of the protein; the repeats could contain specific compositional elements that promote prion propagation; or the repeats, while not essential for prion propagation, might explain some unique features of [PSI(+)]. Here, we test these three hypotheses and show that the ability of the Sup35 and PrP repeats to support [PSI(+)] propagation stems from their amino acid composition, not their primary sequences. Furthermore, we demonstrate that compositional requirements for the repeat domain are distinct from those of the nucleation domain, indicating that prion nucleation and propagation are driven by distinct compositional features.
[PSI(+)],酵母 Sup35 蛋白的朊病毒形式,是由 Sup35 从可溶性形式结构转化为传染性淀粉样形式而产生的。朊病毒的感染力被认为源自伴侣蛋白依赖的纤维切割,这种切割将大的朊病毒纤维断裂成更小的、可遗传的传播体。与哺乳动物朊病毒蛋白 PrP 一样,Sup35 含有寡肽重复结构域。缺失分析表明,寡肽重复结构域对 [PSI(+)]的传播至关重要,而朊病毒结构域的一个独特区域负责朊病毒的成核。PrP 的寡肽重复结构域可以替代 Sup35 的寡肽重复结构域来支持 [PSI(+)]的传播,这表明重复在支持朊病毒维持方面发挥着共同的作用。然而,随机改变 Sup35 朊病毒结构域中氨基酸的顺序并不能阻止朊病毒的形成或传播,这表明氨基酸组成是 Sup35 朊病毒倾向的主要决定因素。因此,寡肽重复在 [PSI(+)]传播中的作用尚不清楚:重复可能只是作为一个非特异性间隔物,将朊病毒成核结构域与蛋白质的其余部分分隔开来;重复可能包含促进朊病毒传播的特定组成元素;或者重复虽然对朊病毒的传播不是必需的,但可能解释了 [PSI(+)]的一些独特特征。在这里,我们检验了这三个假设,并表明 Sup35 和 PrP 重复序列支持 [PSI(+)]传播的能力源于它们的氨基酸组成,而不是它们的原始序列。此外,我们证明了重复结构域的组成要求与成核结构域的要求不同,这表明朊病毒的成核和传播是由不同的组成特征驱动的。