Department of Biosciences, University of Exeter, Exeter EX4 4QD, United Kingdom.
Mol Biol Cell. 2011 Oct;22(19):3645-57. doi: 10.1091/mbc.E11-03-0217. Epub 2011 Aug 10.
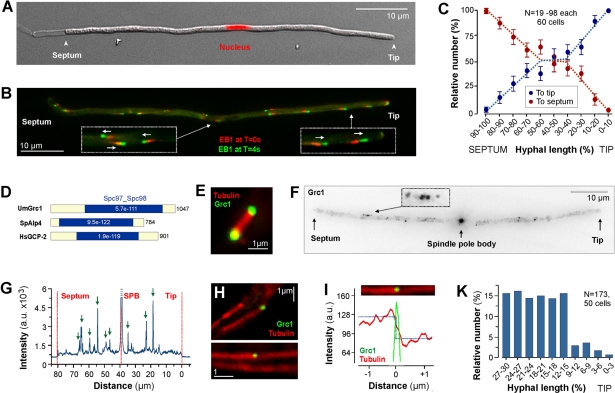
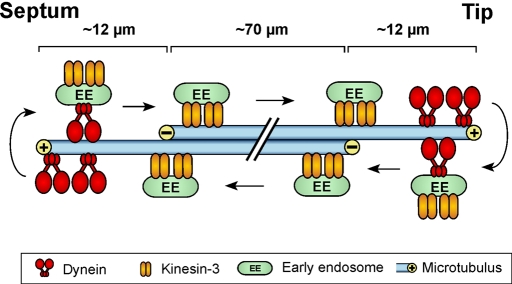
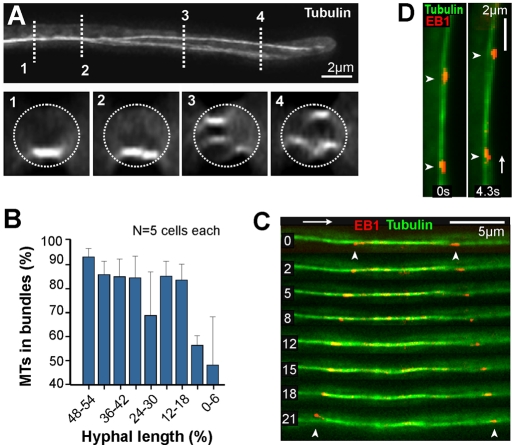
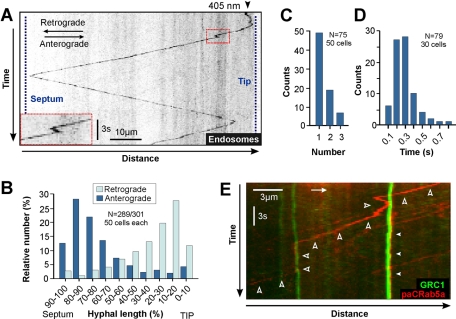
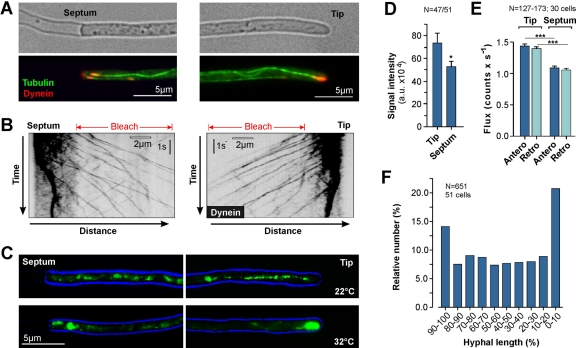
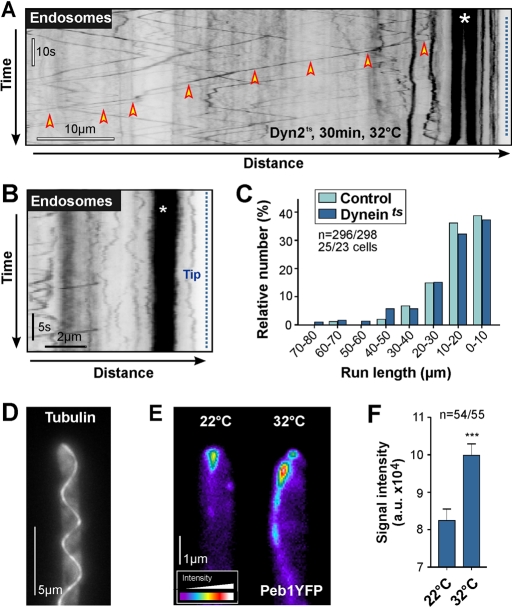
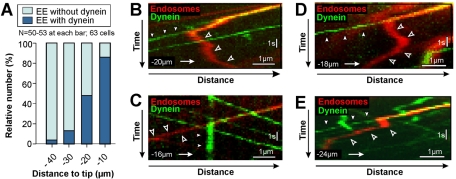
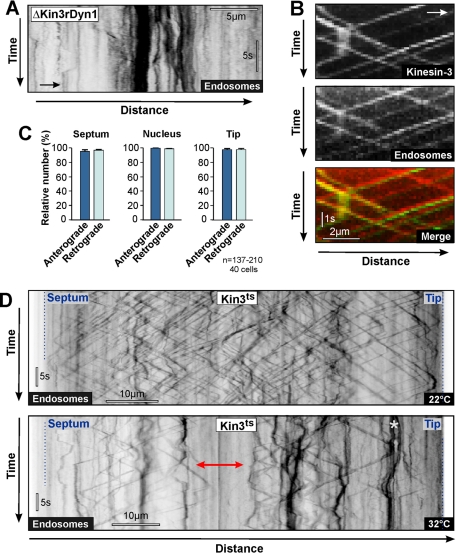
The polarity of microtubules (MTs) determines the motors for intracellular motility, with kinesins moving to plus ends and dynein to minus ends. In elongated cells of Ustilago maydis, dynein is thought to move early endosomes (EEs) toward the septum (retrograde), whereas kinesin-3 transports them to the growing cell tip (anterograde). Occasionally, EEs run up to 90 μm in one direction. The underlying MT array consists of unipolar MTs at both cell ends and antipolar bundles in the middle region of the cell. Cytoplasmic MT-organizing centers, labeled with a γ-tubulin ring complex protein, are distributed along the antipolar MTs but are absent from the unipolar regions. Dynein colocalizes with EEs for 10-20 μm after they have left the cell tip. Inactivation of temperature-sensitive dynein abolishes EE motility within the unipolar MT array, whereas long-range motility is not impaired. In contrast, kinesin-3 is continuously present, and its inactivation stops long-range EE motility. This indicates that both motors participate in EE motility, with dynein transporting the organelles through the unipolar MT array near the cell ends, and kinesin-3 taking over at the beginning of the medial antipolar MT array. The cooperation of both motors mediates EE movements over the length of the entire cell.
微管(MTs)的极性决定了细胞内运动的马达,其中驱动蛋白向正极移动,而动力蛋白向负极移动。在玉米黑粉菌的伸长细胞中,动力蛋白被认为将早期内体(EEs)向隔膜(逆行)移动,而驱动蛋白-3 将它们运输到生长中的细胞尖端(顺行)。偶尔,EEs 在一个方向上可以运行长达 90 μm。下面的微管阵列由细胞两端的单极 MTs 和细胞中部的反极束组成。细胞质微管组织中心,用γ-微管蛋白环复合物蛋白标记,沿着反极微管分布,但不存在于单极区域。在 EEs 离开细胞尖端后 10-20 μm 处,动力蛋白与 EEs 共定位。温度敏感型动力蛋白失活会破坏单极 MT 阵列内 EEs 的运动,但不会损害长程运动。相反,驱动蛋白-3 持续存在,其失活会停止长程 EE 运动。这表明两种马达都参与了 EE 的运动,动力蛋白通过细胞末端附近的单极 MT 阵列运输细胞器,而驱动蛋白-3 在中部反极 MT 阵列的开始时接管。两种马达的协同作用介导了 EE 在整个细胞长度上的运动。