National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, MD 20894, USA.
Nucleic Acids Res. 2011 Dec;39(22):9473-97. doi: 10.1093/nar/gkr691. Epub 2011 Sep 3.
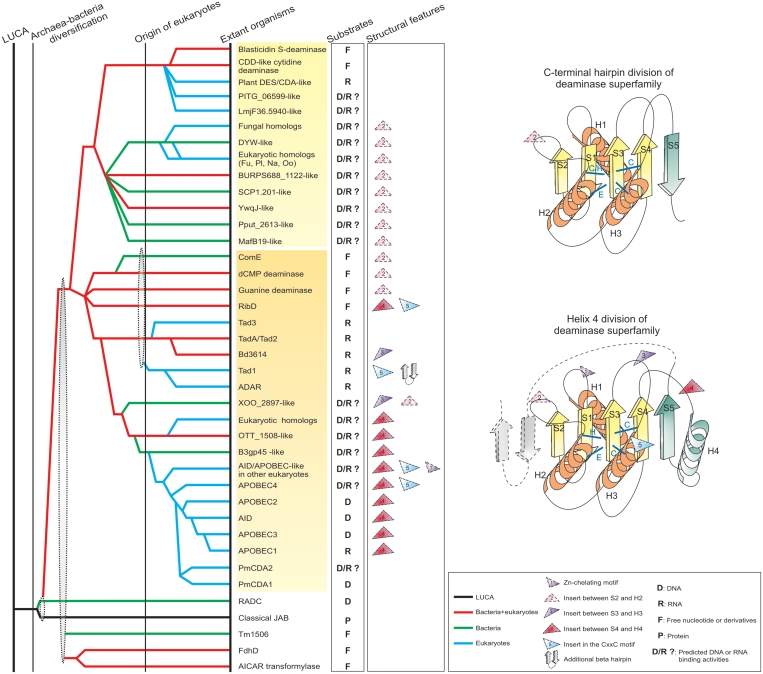
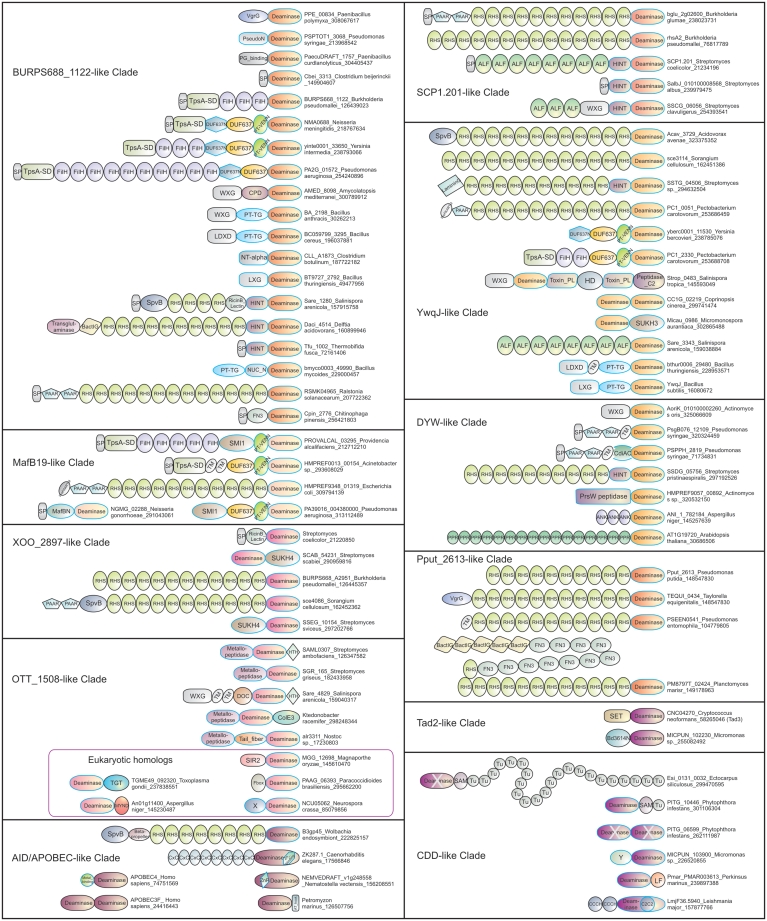
The deaminase-like fold includes, in addition to nucleic acid/nucleotide deaminases, several catalytic domains such as the JAB domain, and others involved in nucleotide and ADP-ribose metabolism. Using sensitive sequence and structural comparison methods, we develop a comprehensive natural classification of the deaminase-like fold and show that its ancestral version was likely to operate on nucleotides or nucleic acids. Consequently, we present evidence that a specific group of JAB domains are likely to possess a DNA repair function, distinct from the previously known deubiquitinating peptidase activity. We also identified numerous previously unknown clades of nucleic acid deaminases. Using inference based on contextual information, we suggest that most of these clades are toxin domains of two distinct classes of bacterial toxin systems, namely polymorphic toxins implicated in bacterial interstrain competition and those that target distantly related cells. Genome context information suggests that these toxins might be delivered via diverse secretory systems, such as Type V, Type VI, PVC and a novel PrsW-like intramembrane peptidase-dependent mechanism. We propose that certain deaminase toxins might be deployed by diverse extracellular and intracellular pathogens as also endosymbionts as effectors targeting nucleic acids of host cells. Our analysis suggests that these toxin deaminases have been acquired by eukaryotes on several independent occasions and recruited as organellar or nucleo-cytoplasmic RNA modifiers, operating on tRNAs, mRNAs and short non-coding RNAs, and also as mutators of hyper-variable genes, viruses and selfish elements. This scenario potentially explains the origin of mutagenic AID/APOBEC-like deaminases, including novel versions from Caenorhabditis, Nematostella and diverse algae and a large class of fast-evolving fungal deaminases. These observations greatly expand the distribution of possible unidentified mutagenic processes catalyzed by nucleic acid deaminases.
脱氨酶样折叠结构除了包含核酸/核苷酸脱氨酶以外,还有一些催化结构域,如 JAB 结构域,以及其他参与核苷酸和 ADP-核糖代谢的结构域。通过使用灵敏的序列和结构比较方法,我们对脱氨酶样折叠结构进行了全面的自然分类,并表明其原始版本可能作用于核苷酸或核酸。因此,我们提出了证据,表明一组特定的 JAB 结构域可能具有 DNA 修复功能,与先前已知的去泛素化肽酶活性不同。我们还鉴定了许多以前未知的核酸脱氨酶类群。通过基于上下文信息的推断,我们认为这些类群中的大多数是两种不同类型的细菌毒素系统的毒素结构域,即参与细菌种间竞争的多态毒素和针对远缘细胞的毒素。基因组上下文信息表明,这些毒素可能通过多种分泌系统传递,如类型 V、类型 VI、PVC 和一种新的 PrsW 样内膜肽酶依赖机制。我们提出,某些脱氨酶毒素可能被多种细胞外和细胞内病原体以及内共生体作为针对宿主细胞核酸的效应物来部署。我们的分析表明,这些毒素脱氨酶已在几个独立的场合被真核生物获得,并作为细胞器或核质 RNA 修饰物被招募,作用于 tRNA、mRNA 和短非编码 RNA,以及作为高变基因、病毒和自私元件的突变剂。这种情况可能解释了 AID/APOBEC 样脱氨酶的起源,包括来自秀丽隐杆线虫、海葵和多种藻类的新型版本以及一大类快速进化的真菌脱氨酶。这些观察结果大大扩展了可能由核酸脱氨酶催化的未知诱变过程的分布。