National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, MD 20894, USA.
Biol Direct. 2012 Jun 25;7:18. doi: 10.1186/1745-6150-7-18.
Proteinaceous toxins are observed across all levels of inter-organismal and intra-genomic conflicts. These include recently discovered prokaryotic polymorphic toxin systems implicated in intra-specific conflicts. They are characterized by a remarkable diversity of C-terminal toxin domains generated by recombination with standalone toxin-coding cassettes. Prior analysis revealed a striking diversity of nuclease and deaminase domains among the toxin modules. We systematically investigated polymorphic toxin systems using comparative genomics, sequence and structure analysis.
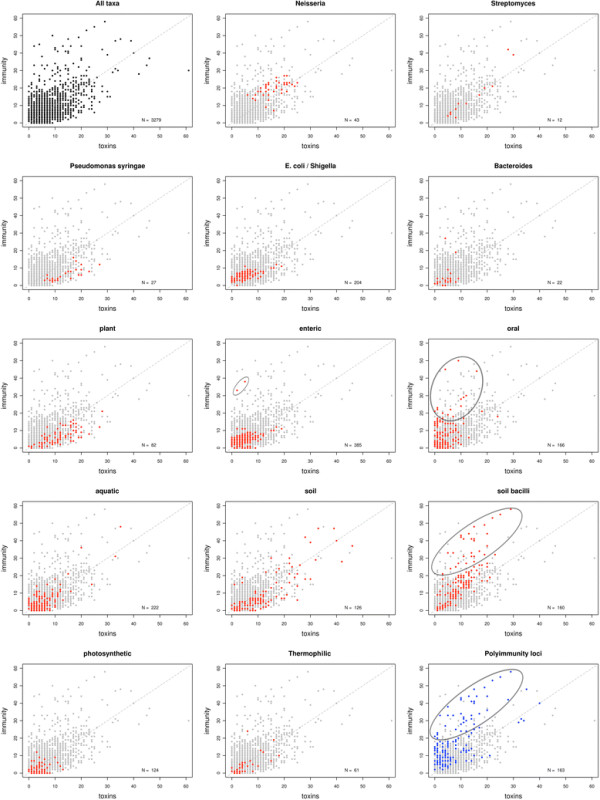
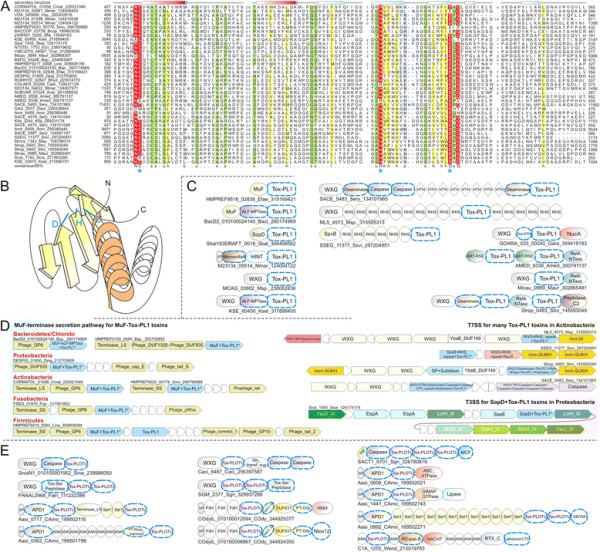
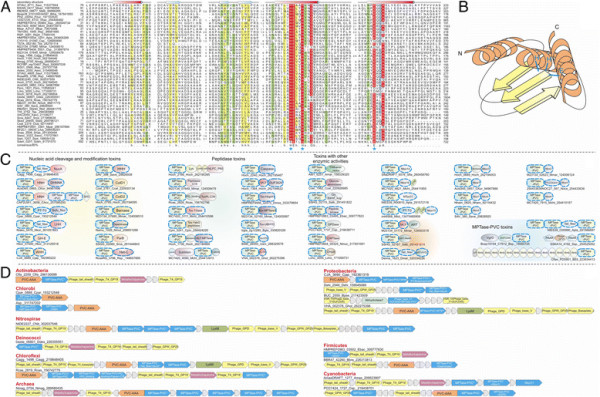
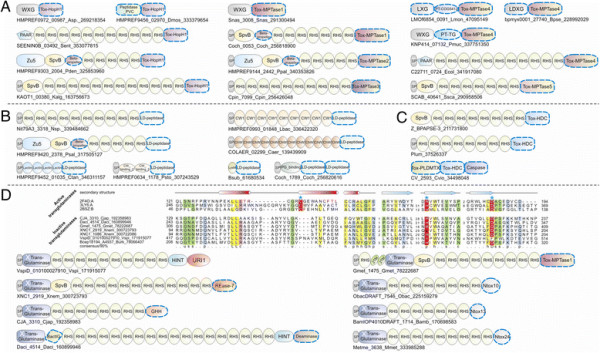
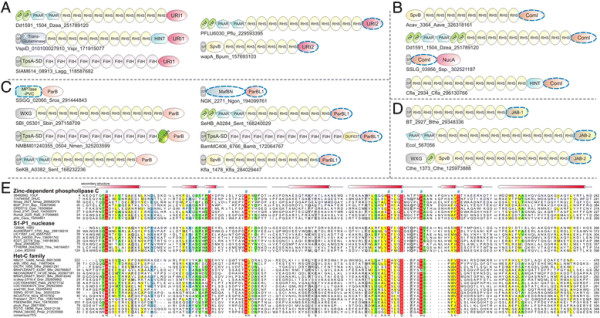
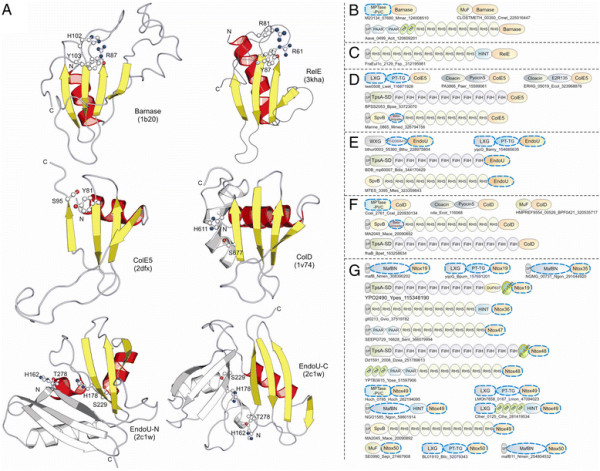
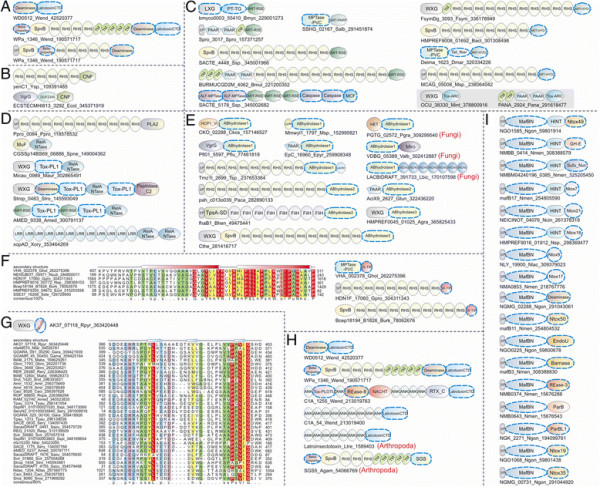
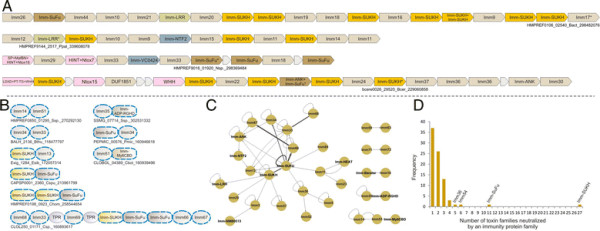
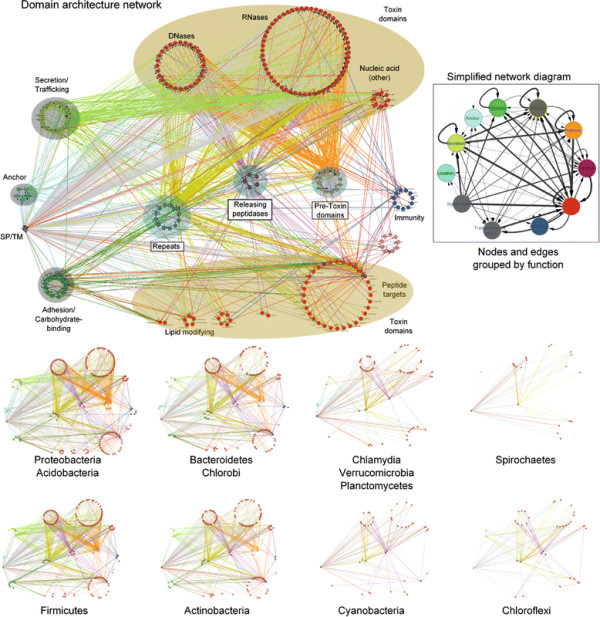
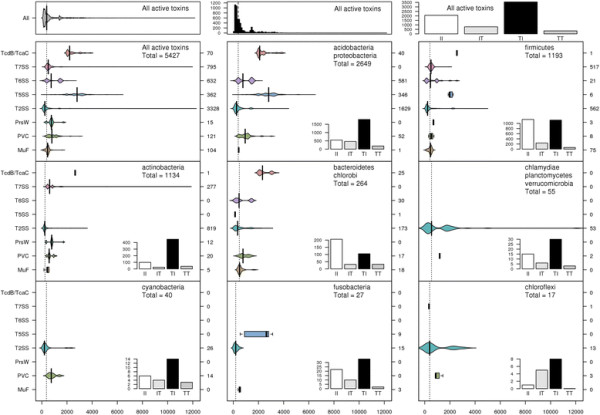
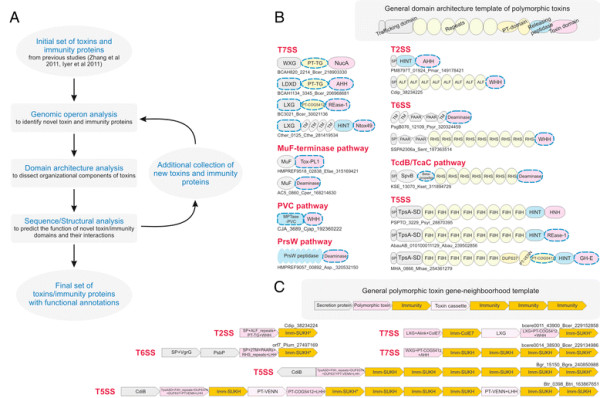
Polymorphic toxin systems are distributed across all major bacterial lineages and are delivered by at least eight distinct secretory systems. In addition to type-II, these include type-V, VI, VII (ESX), and the poorly characterized "Photorhabdus virulence cassettes (PVC)", PrsW-dependent and MuF phage-capsid-like systems. We present evidence that trafficking of these toxins is often accompanied by autoproteolytic processing catalyzed by HINT, ZU5, PrsW, caspase-like, papain-like, and a novel metallopeptidase associated with the PVC system. We identified over 150 distinct toxin domains in these systems. These span an extraordinary catalytic spectrum to include 23 distinct clades of peptidases, numerous previously unrecognized versions of nucleases and deaminases, ADP-ribosyltransferases, ADP ribosyl cyclases, RelA/SpoT-like nucleotidyltransferases, glycosyltranferases and other enzymes predicted to modify lipids and carbohydrates, and a pore-forming toxin domain. Several of these toxin domains are shared with host-directed effectors of pathogenic bacteria. Over 90 families of immunity proteins might neutralize anywhere between a single to at least 27 distinct types of toxin domains. In some organisms multiple tandem immunity genes or immunity protein domains are organized into polyimmunity loci or polyimmunity proteins. Gene-neighborhood-analysis of polymorphic toxin systems predicts the presence of novel trafficking-related components, and also the organizational logic that allows toxin diversification through recombination. Domain architecture and protein-length analysis revealed that these toxins might be deployed as secreted factors, through directed injection, or via inter-cellular contact facilitated by filamentous structures formed by RHS/YD, filamentous hemagglutinin and other repeats. Phyletic pattern and life-style analysis indicate that polymorphic toxins and polyimmunity loci participate in cooperative behavior and facultative 'cheating' in several ecosystems such as the human oral cavity and soil. Multiple domains from these systems have also been repeatedly transferred to eukaryotes and their viruses, such as the nucleo-cytoplasmic large DNA viruses.
Along with a comprehensive inventory of toxins and immunity proteins, we present several testable predictions regarding active sites and catalytic mechanisms of toxins, their processing and trafficking and their role in intra-specific and inter-specific interactions between bacteria. These systems provide insights regarding the emergence of key systems at different points in eukaryotic evolution, such as ADP ribosylation, interaction of myosin VI with cargo proteins, mediation of apoptosis, hyphal heteroincompatibility, hedgehog signaling, arthropod toxins, cell-cell interaction molecules like teneurins and different signaling messengers.
蛋白毒素存在于生物组织间和基因组内的所有冲突级别中。其中包括最近发现的与种内冲突有关的原核多态性毒素系统。这些系统的特点是通过与独立的毒素编码盒重组产生了显著多样化的 C 端毒素结构域。先前的分析显示,毒素模块中存在着惊人多样的核酸酶和脱氨酶结构域。我们使用比较基因组学、序列和结构分析系统地研究了多态性毒素系统。
多态性毒素系统分布在所有主要的细菌谱系中,并通过至少八种不同的分泌系统传递。除了 II 型之外,这些系统还包括 V、VI、VII(ESX)和特征不明显的“Photorhabdus 毒力盒(PVC)”、PrsW 依赖性和 MuF 噬菌体衣壳样系统。我们提供的证据表明,这些毒素的运输通常伴随着由 HINT、ZU5、PrsW、半胱氨酸蛋白酶样、木瓜蛋白酶样和与 PVC 系统相关的新型金属肽酶催化的自蛋白水解加工。我们在这些系统中鉴定了 150 多种不同的毒素结构域。这些结构域跨越了非凡的催化谱,包括 23 个不同的肽酶簇、许多以前未被识别的核酸酶和脱氨酶、ADP-核糖基转移酶、ADP 核糖基环化酶、RelA/SpoT 样核苷酸转移酶、糖基转移酶以及其他预测能够修饰脂质和碳水化合物的酶,以及一个形成孔的毒素结构域。这些毒素结构域中的一些与致病性细菌的宿主定向效应物共享。超过 90 种免疫蛋白家族可能中和单一至至少 27 种不同类型的毒素结构域。在某些生物体中,多个串联免疫基因或免疫蛋白结构域被组织成多免疫基因座或多免疫蛋白。多态性毒素系统的基因邻域分析预测了新型运输相关成分的存在,以及通过重组产生毒素多样化的组织逻辑。结构域架构和蛋白质长度分析表明,这些毒素可能作为分泌因子被部署,通过定向注射,或通过由 RHS/YD、丝状血凝素和其他重复序列形成的丝状结构通过细胞间接触进行。系统发生模式和生活方式分析表明,多态性毒素和多免疫基因座参与了几种生态系统中的合作行为和兼性“欺骗”,例如人类口腔和土壤。这些系统的多个结构域也被反复转移到真核生物及其病毒中,如核质大 DNA 病毒。
除了对毒素和免疫蛋白进行全面盘点外,我们还提出了一些关于毒素、它们的加工和运输的活性位点和催化机制以及它们在细菌种内和种间相互作用中的作用的可测试预测。这些系统为研究真核生物进化过程中的关键系统提供了线索,例如 ADP 核糖基化、肌球蛋白 VI 与货物蛋白的相互作用、细胞凋亡的介导、菌丝异质相容性、 hedgehog 信号转导、节肢动物毒素、细胞-细胞相互作用分子如 tenurin 和不同的信号信使。