Neuroscience Group, Institute for Science and Technology in Medicine, Keele University, Keele, United Kingdom.
PLoS One. 2011;6(9):e24892. doi: 10.1371/journal.pone.0024892. Epub 2011 Sep 15.
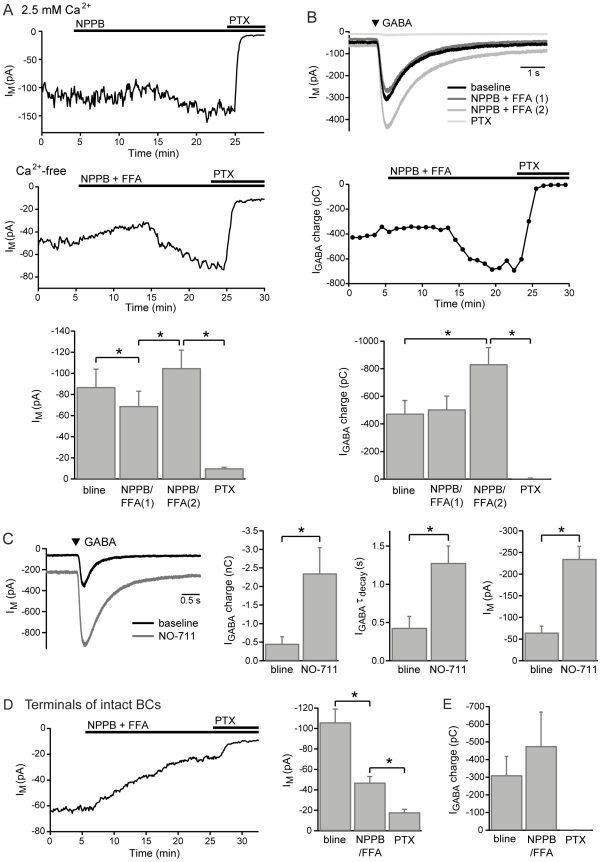
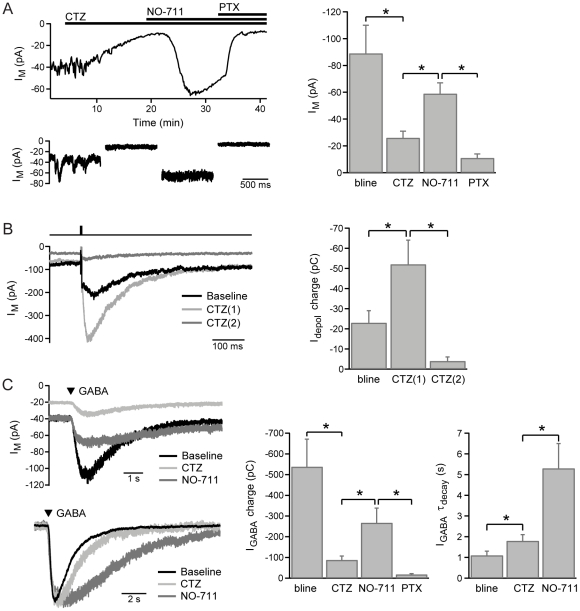
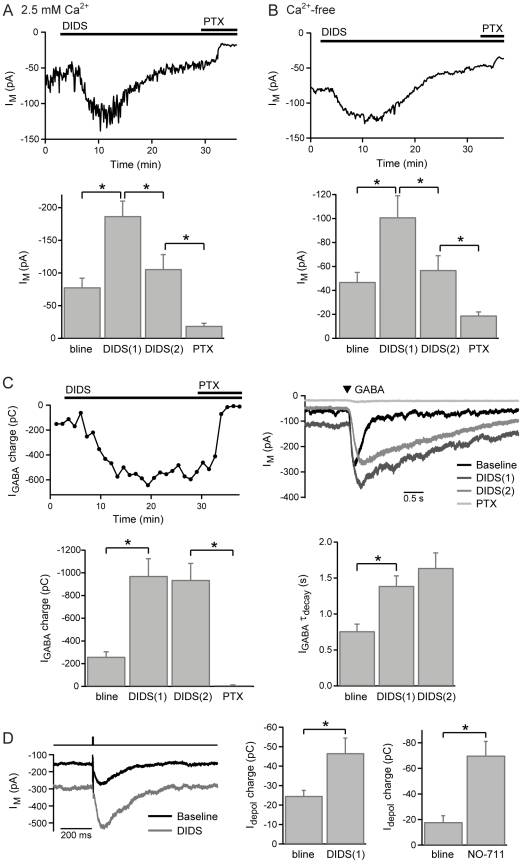
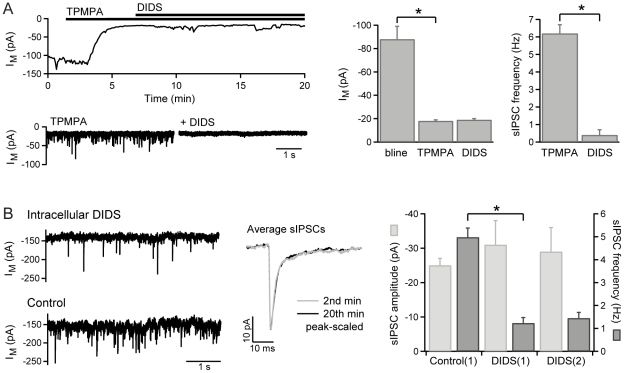
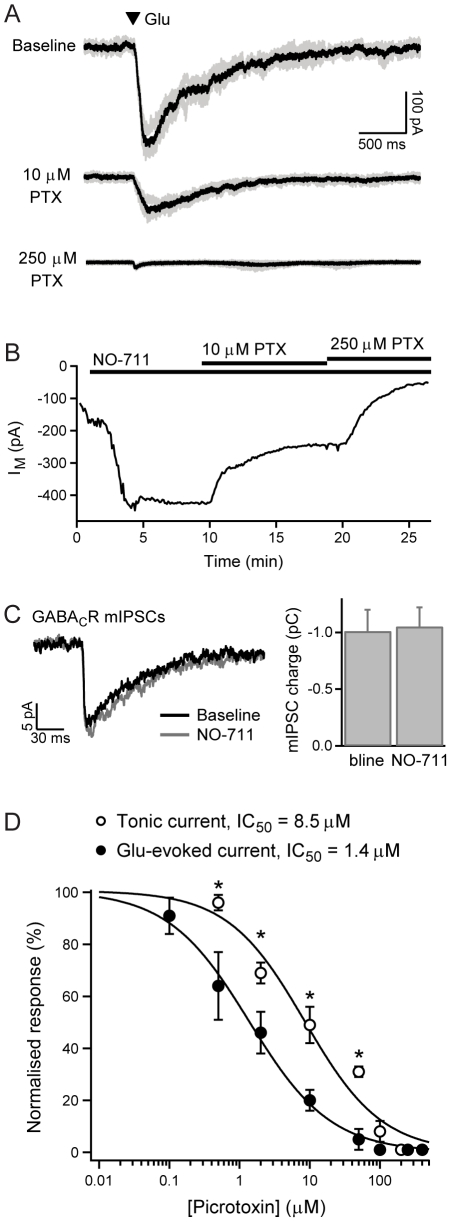
GABAergic inhibition in the central nervous system (CNS) can occur via rapid, transient postsynaptic currents and via a tonic increase in membrane conductance, mediated by synaptic and extrasynaptic GABA(A) receptors (GABA(A)Rs) respectively. Retinal bipolar cells (BCs) exhibit a tonic current mediated by GABA(C)Rs in their axon terminal, in addition to synaptic GABA(A)R and GABA(C)R currents, which strongly regulate BC output. The tonic GABA(C)R current in BC terminals (BCTs) is not dependent on vesicular GABA release, but properties such as the alternative source of GABA and the identity of the GABA(C)Rs remain unknown. Following a recent report that tonic GABA release from cerebellar glial cells is mediated by Bestrophin 1 anion channels, we have investigated their role in non-vesicular GABA release in the retina. Using patch-clamp recordings from BCTs in goldfish retinal slices, we find that the tonic GABA(C)R current is not reduced by the anion channel inhibitors NPPB or flufenamic acid but is reduced by DIDS, which decreases the tonic current without directly affecting GABA(C)Rs. All three drugs also exhibit non-specific effects including inhibition of GABA transporters. GABA(C)R ρ subunits can form homomeric and heteromeric receptors that differ in their properties, but BC GABA(C)Rs are thought to be ρ1-ρ2 heteromers. To investigate whether GABA(C)Rs mediating tonic and synaptic currents may differ in their subunit composition, as is the case for GABA(A)Rs, we have examined the effects of two antagonists that show partial ρ subunit selectivity: picrotoxin and cyclothiazide. Tonic and synaptic GABA(C)R currents were differentially affected by both drugs, suggesting that a population of homomeric ρ1 receptors contributes to the tonic current. These results extend our understanding of the multiple forms of GABAergic inhibition that exist in the CNS and contribute to visual signal processing in the retina.
中枢神经系统(CNS)中的 GABA 能抑制可以通过快速、短暂的突触后电流和通过突触和 extrasynaptic GABA(A) 受体(GABA(A)Rs)介导的膜电导的持续增加来发生。视网膜双极细胞(BCs)在其轴突末梢表现出由 GABA(C)Rs 介导的持续电流,除了突触 GABA(A)R 和 GABA(C)R 电流外,这些电流强烈调节 BC 输出。BC 末梢(BCTs)中的持续 GABA(C)R 电流不依赖于囊泡 GABA 释放,但 GABA 的替代来源和 GABA(C)Rs 的身份等特性仍不清楚。在最近的一份报告称小脑胶质细胞的持续 GABA 释放是由 Bestrophin 1 阴离子通道介导的之后,我们研究了它们在视网膜中非囊泡 GABA 释放中的作用。我们使用金鱼视网膜切片中的 BCT 进行膜片钳记录,发现阴离子通道抑制剂 NPPB 或 flufenamic acid 并未减少持续的 GABA(C)R 电流,但 DIDS 减少了持续电流,而不会直接影响 GABA(C)Rs。这三种药物还表现出非特异性作用,包括抑制 GABA 转运体。GABA(C)R ρ 亚基可以形成具有不同特性的同源和异源受体,但 BC GABA(C)Rs 被认为是 ρ1-ρ2 异源受体。为了研究调节持续和突触电流的 GABA(C)Rs 是否在亚基组成上可能不同,就像 GABA(A)Rs 一样,我们检查了两种显示部分 ρ 亚基选择性的拮抗剂的影响:荷包牡丹碱和环噻嗪。两种药物都对持续和突触 GABA(C)R 电流有不同的影响,表明同源 ρ1 受体群体有助于持续电流。这些结果扩展了我们对中枢神经系统中存在的多种 GABA 能抑制形式的理解,并有助于视网膜中的视觉信号处理。