The Institute of Structural and Molecular Biology, University of Edinburgh, Edinburgh, United Kingdom.
PLoS One. 2012;7(4):e34820. doi: 10.1371/journal.pone.0034820. Epub 2012 Apr 10.
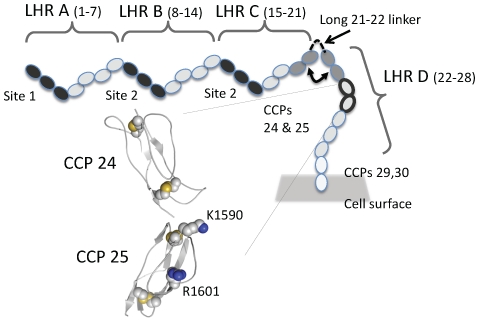
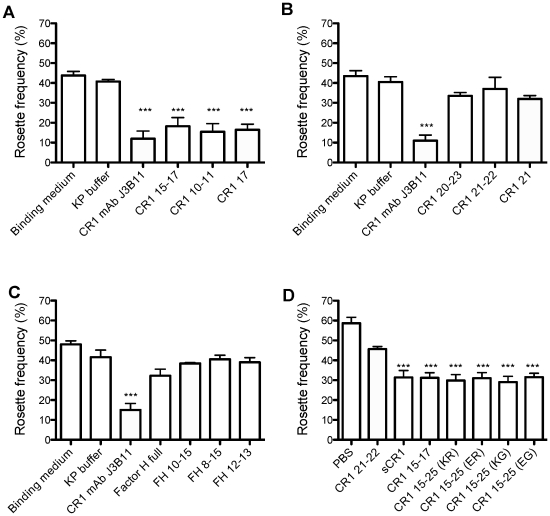
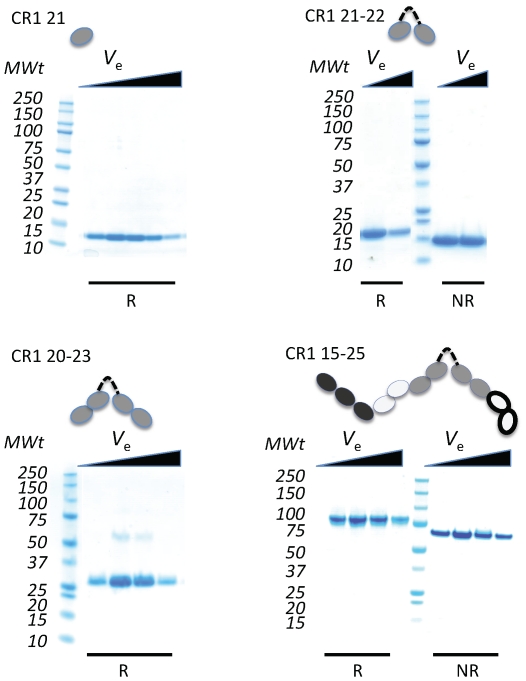
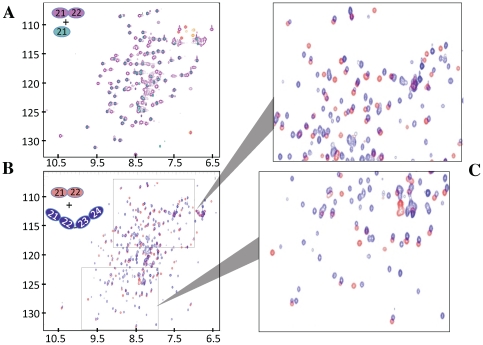
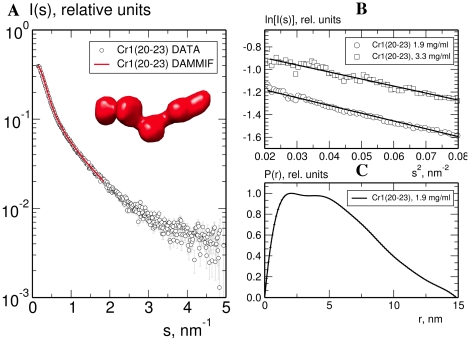
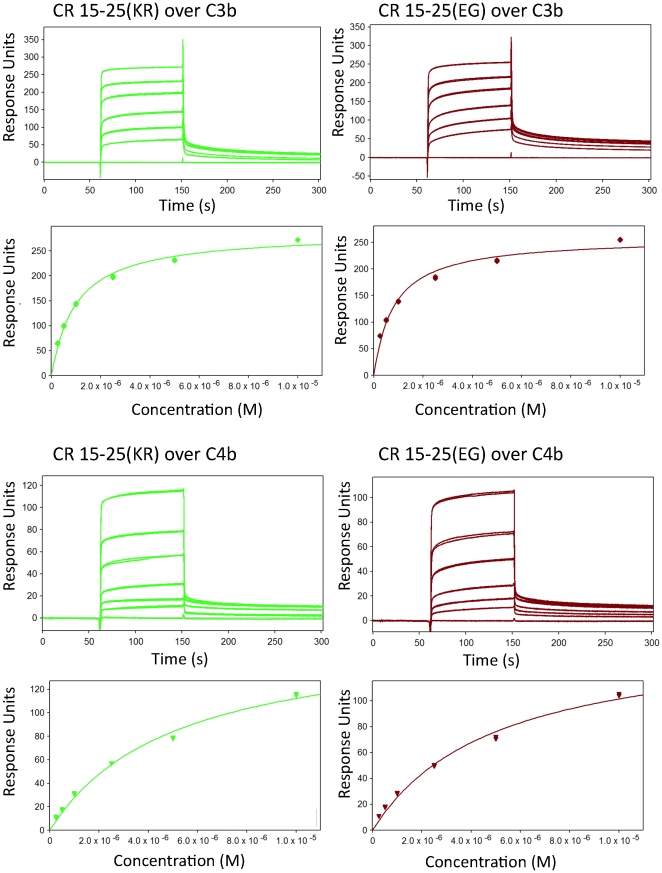
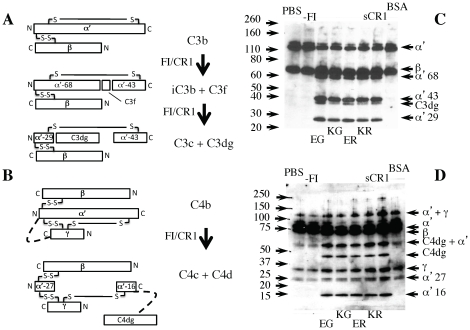
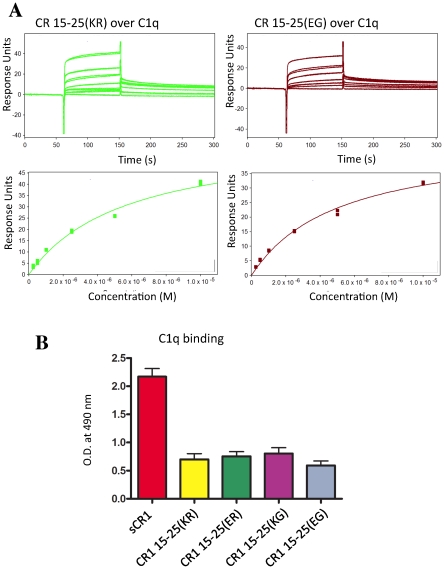
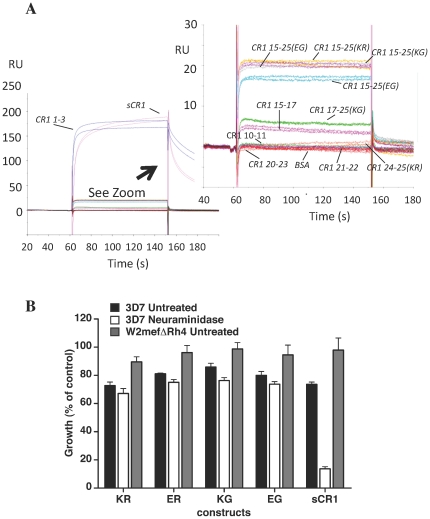
Complement receptor-type 1 (CR1, CD35) is the immune-adherence receptor, a complement regulator, and an erythroid receptor for Plasmodium falciparum during merozoite invasion and subsequent rosette formation involving parasitized and non-infected erythrocytes. The non-uniform geographical distribution of Knops blood group CR1 alleles Sl1/2 and McC(a/b) may result from selective pressures exerted by differential exposure to infectious hazards. Here, four variant short recombinant versions of CR1 were produced and analyzed, focusing on complement control protein modules (CCPs) 15-25 of its ectodomain. These eleven modules encompass a region (CCPs 15-17) key to rosetting, opsonin recognition and complement regulation, as well as the Knops blood group polymorphisms in CCPs 24-25. All four CR1 15-25 variants were monomeric and had similar axial ratios. Modules 21 and 22, despite their double-length inter-modular linker, did not lie side-by-side so as to stabilize a bent-back architecture that would facilitate cooperation between key functional modules and Knops blood group antigens. Indeed, the four CR1 15-25 variants had virtually indistinguishable affinities for immobilized complement fragments C3b (K(D) = 0.8-1.1 µM) and C4b (K(D) = 5.0-5.3 µM). They were all equally good co-factors for factor I-catalysed cleavage of C3b and C4b, and they bound equally within a narrow affinity range, to immobilized C1q. No differences between the variants were observed in assays for inhibition of erythrocyte invasion by P. falciparum or for rosette disruption. Neither differences in complement-regulatory functionality, nor interactions with P. falciparum proteins tested here, appear to have driven the non-uniform geographic distribution of these alleles.
补体受体 1 型(CR1,CD35)是免疫粘附受体、补体调节剂和疟原虫裂殖子入侵和随后涉及寄生和未感染红细胞的玫瑰花形成的红细胞受体。Knops 血型 CR1 等位基因 Sl1/2 和 McC(a/b) 的非均匀地理分布可能是由于对不同感染危害的选择性压力造成的。在这里,产生并分析了四个变异的短重组 CR1 版本,重点是其外域的补体控制蛋白模块(CCPs)15-25。这十一个模块包含一个关键的玫瑰花形成区域(CCPs 15-17)、调理素识别和补体调节,以及 CCPs 24-25 中的 Knops 血型多态性。所有四个 CR1 15-25 变体都是单体,具有相似的轴比。尽管模块 21 和 22 具有双长度的模块间接头,但它们并没有并排排列,从而稳定了一个弯曲的后向结构,这将有助于关键功能模块和 Knops 血型抗原之间的合作。事实上,四个 CR1 15-25 变体对固定化补体片段 C3b(K(D)=0.8-1.1 μM)和 C4b(K(D)=5.0-5.3 μM)的亲和力几乎无法区分。它们都是因子 I 催化切割 C3b 和 C4b 的同样良好的辅助因子,并且在狭窄的亲和力范围内,它们都与固定化 C1q 结合。在抑制疟原虫对红细胞的入侵或破坏玫瑰花的测定中,没有观察到变体之间的差异。在这里测试的补体调节功能的差异,以及与疟原虫蛋白的相互作用,似乎都没有导致这些等位基因的非均匀地理分布。