Department of Palaeontology, Institut Royal des Sciences Naturelles de Belgique, Bruxelles, Belgium.
PLoS One. 2012;7(5):e36849. doi: 10.1371/journal.pone.0036849. Epub 2012 May 30.
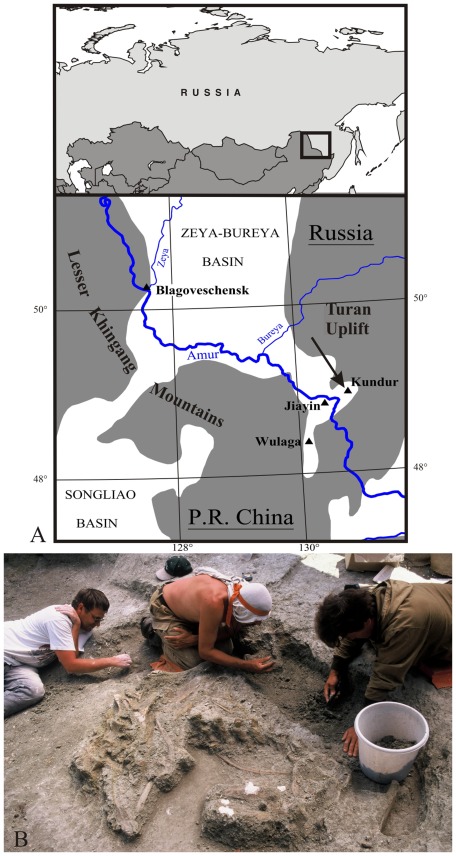
Four main dinosaur sites have been investigated in latest cretaceous deposits from the Amur/Heilongjiang Region: Jiayin and Wulaga in China (Yuliangze Formation), Blagoveschensk and Kundur in Russia (Udurchukan Formation). More than 90% of the bones discovered in these localities belong to hollow-crested lambeosaurine saurolophids, but flat-headed saurolophines are also represented: Kerberosaurus manakini at Blagoveschensk and Wulagasaurus dongi at Wulaga.
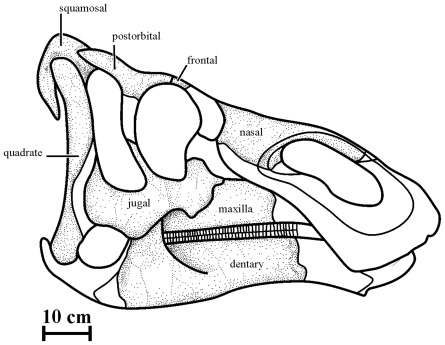
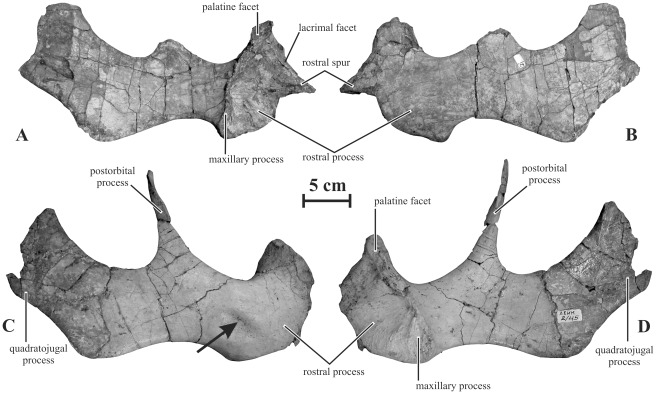
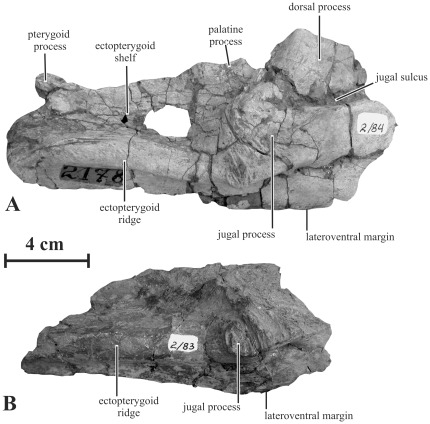
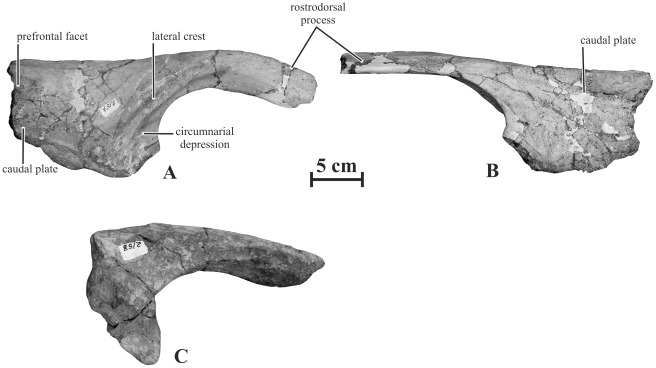
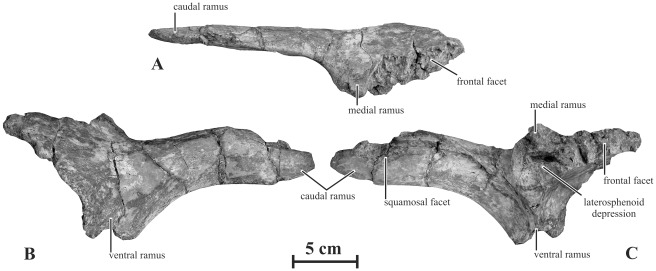
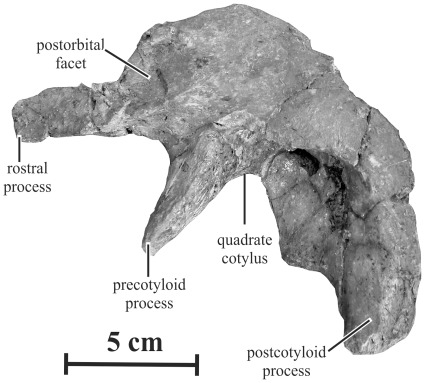
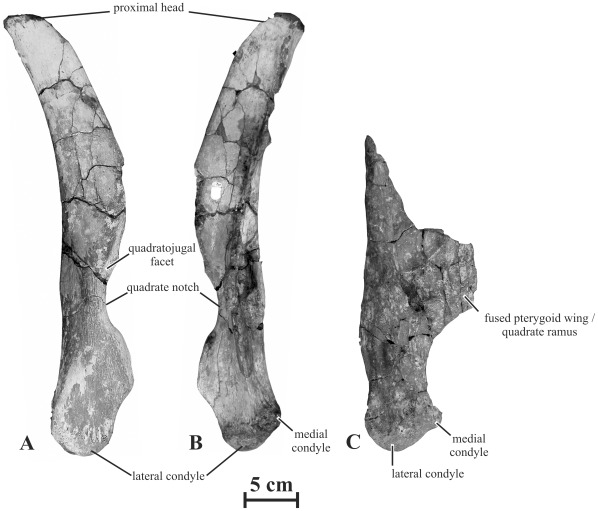
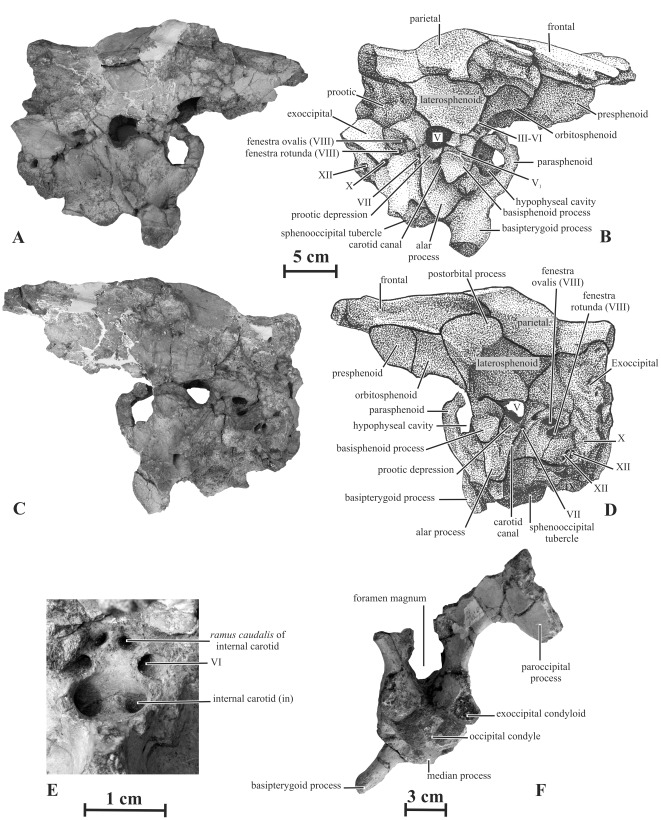
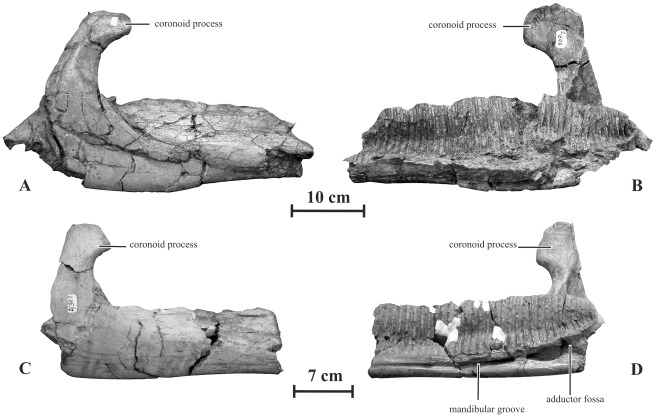
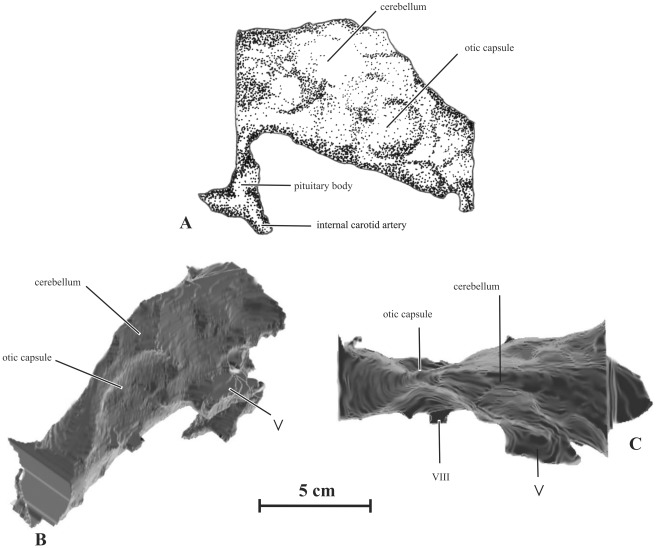
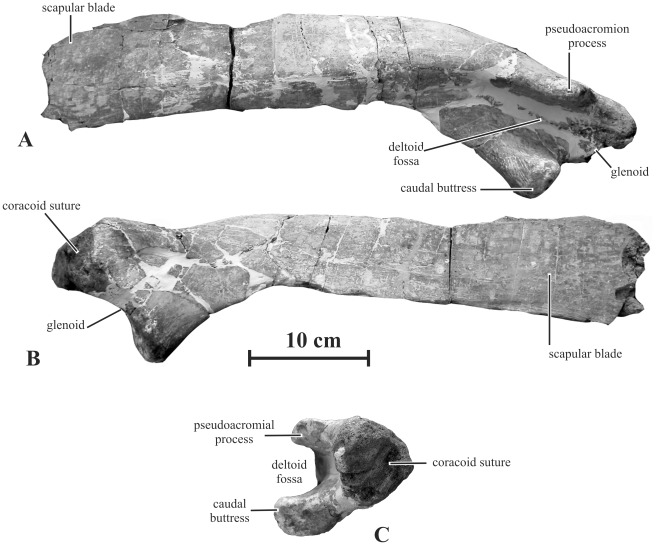
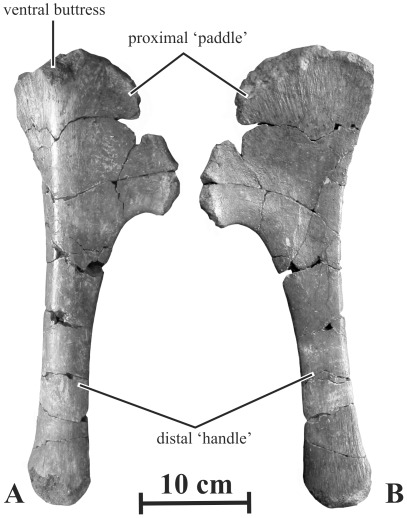
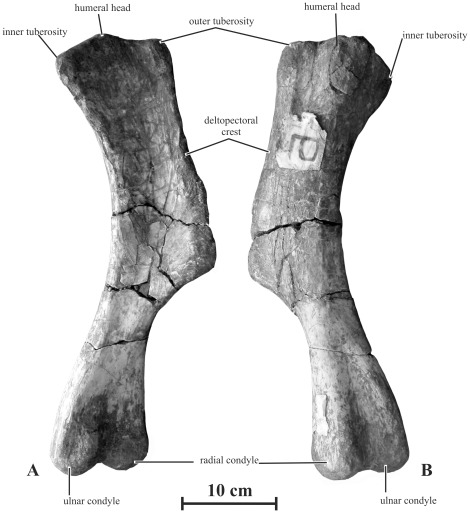
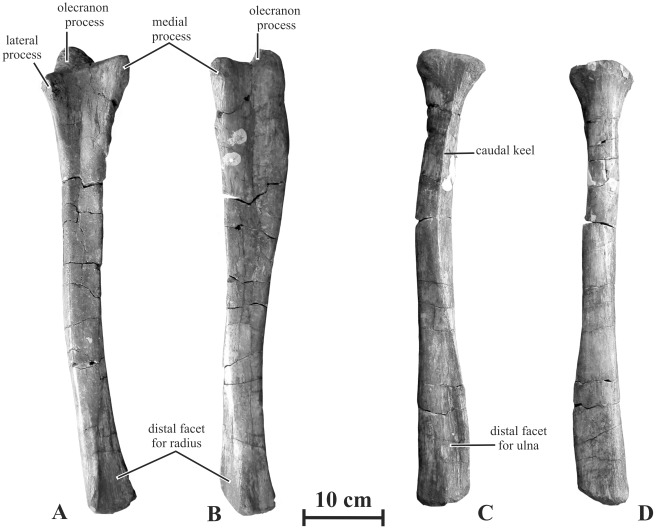
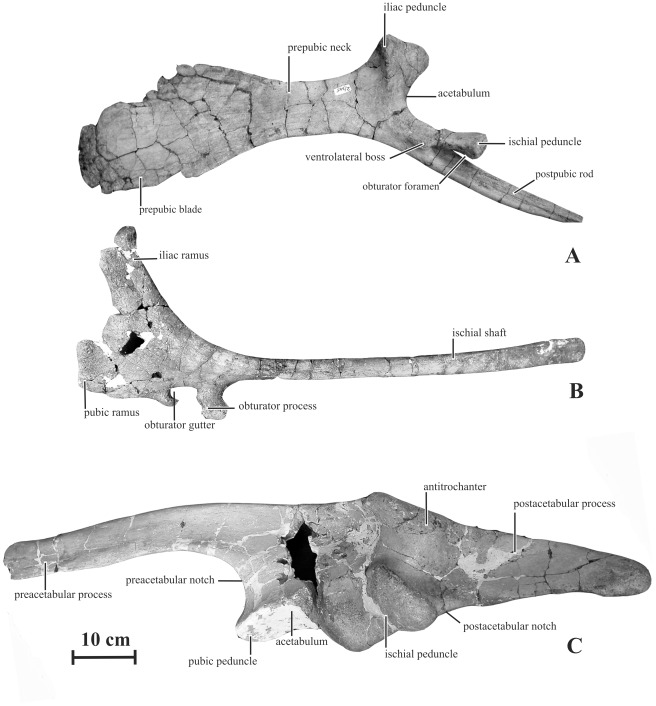
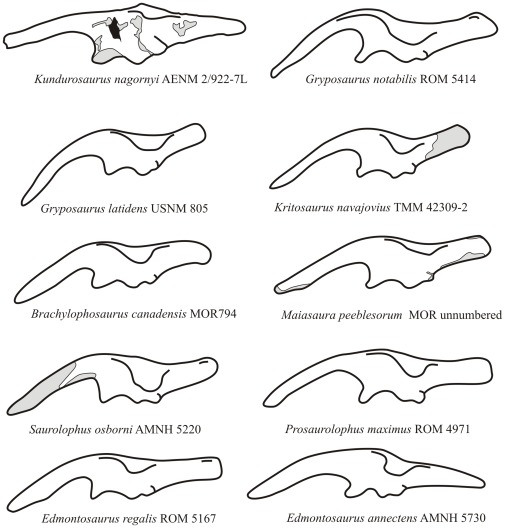
METHODOLOGY/PRINCIPAL FINDINGS: Herein we describe a new saurolophine dinosaur, Kundurosaurus nagornyi gen. et sp. nov., from the Udurchukan Formation (Maastrichtian) of Kundur, represented by disarticulated cranial and postcranial material. This new taxon is diagnosed by four autapomorphies.
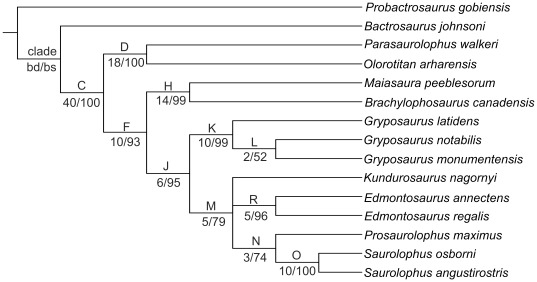
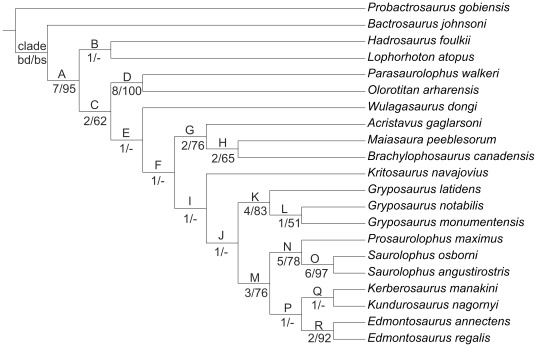
CONCLUSIONS/SIGNIFICANCE: A phylogenetic analysis of saurolophines indicates that Kundurosaurus nagornyi is nested within a rather robust clade including Edmontosaurus spp., Saurolophus spp., and Prosaurolophus maximus, possibly as a sister-taxon for Kerberosaurus manakini also from the Udurchukan Formation of Far Eastern Russia. The high diversity and mosaic distribution of Maastrichtian hadrosaurid faunas in the Amur-Heilongjiang region are the result of a complex palaeogeographical history and imply that many independent hadrosaurid lineages dispersed without any problem between western America and eastern Asia at the end of the Cretaceous.
在黑龙江/阿穆尔地区最新白垩纪沉积物中已经调查了四个主要的恐龙遗址:中国的嘉荫和乌尔加(于亮泽组),俄罗斯的布拉戈维申斯克和坤德尔(乌杜尔恰肯组)。在这些地方发现的骨骼中,超过 90%属于中空冠龙类鸭嘴龙,但平头鸭嘴龙也有代表:布拉戈维申斯克的 Kerberosaurus manakini 和乌尔加的 Wulagasaurus dongi。
方法/主要发现:本文描述了一个来自坤德尔 Udurchukan 组(马斯特里赫特阶)的新的鸭嘴龙类恐龙 Kundurosaurus nagornyi gen. et sp. nov.,由不相连的头骨和后头骨材料代表。这个新的分类单元通过四个独特的特征来诊断。
结论/意义:对鸭嘴龙类的系统发育分析表明,Kundurosaurus nagornyi 嵌套在一个相当强壮的分支中,包括埃德蒙顿龙属、鸭嘴龙属和原鸭嘴龙属,可能与来自俄罗斯远东乌杜尔恰肯组的 Kerberosaurus manakini 是姐妹群。黑龙江/阿穆尔地区马斯特里赫特阶鸭嘴龙类动物群的高度多样性和镶嵌分布是复杂的古地理历史的结果,这意味着许多独立的鸭嘴龙类谱系在白垩纪末期没有任何问题地在美洲西部和亚洲东部之间扩散。