The Lewis-Sigler Institute for Integrative Genomics, Princeton University, Princeton, NJ 08544, USA.
Mol Biol Cell. 2012 Aug;23(15):2993-3007. doi: 10.1091/mbc.E12-03-0232. Epub 2012 Jun 13.
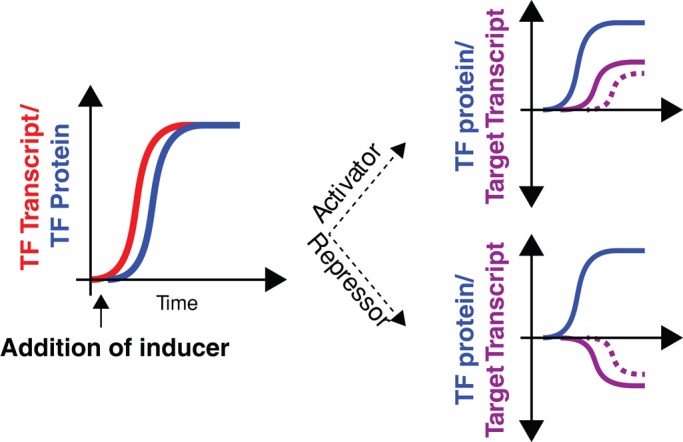
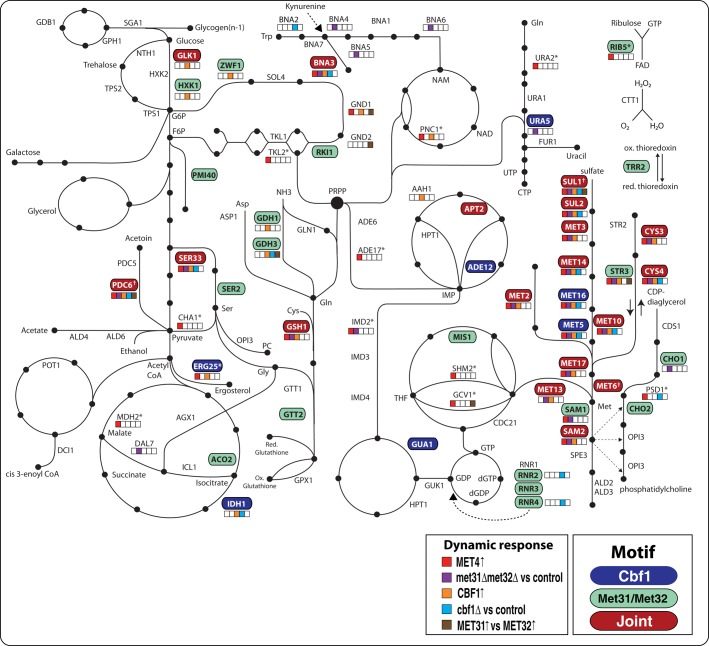
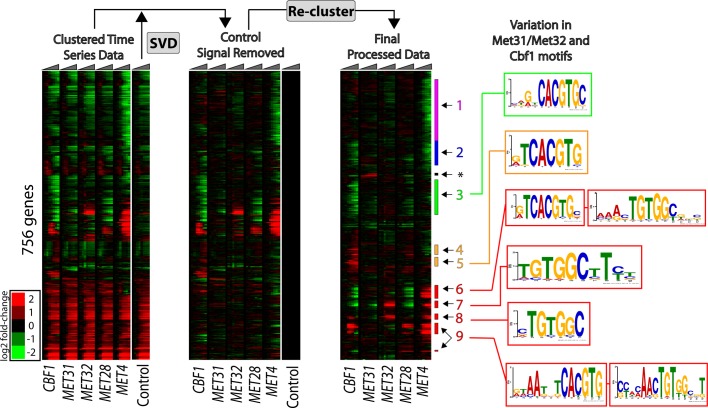
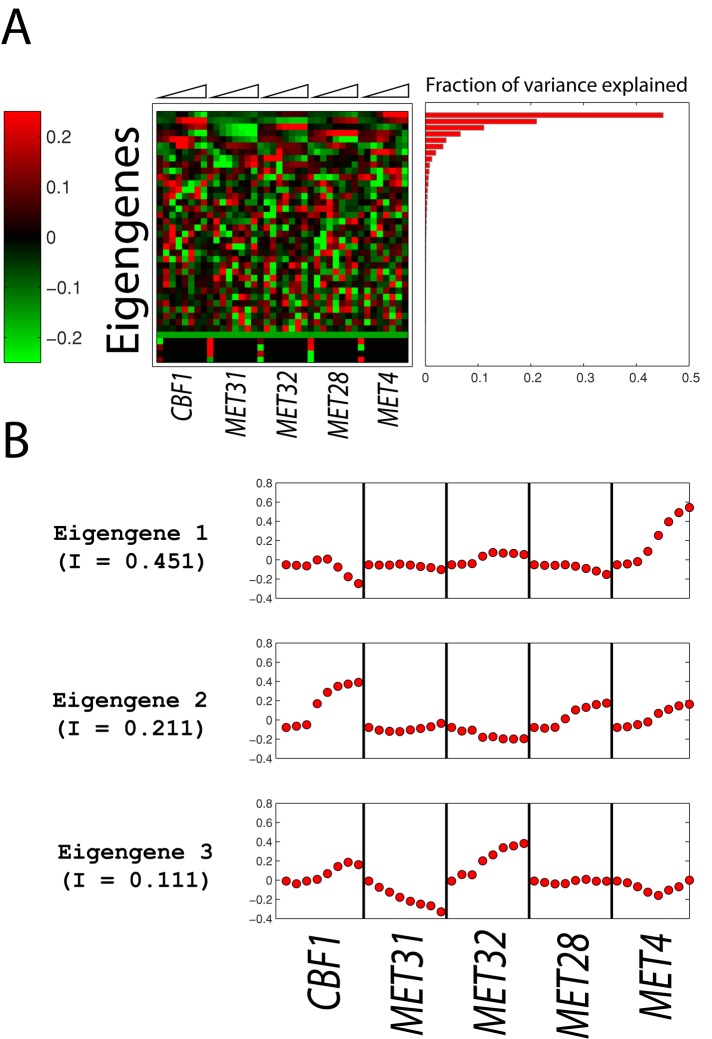
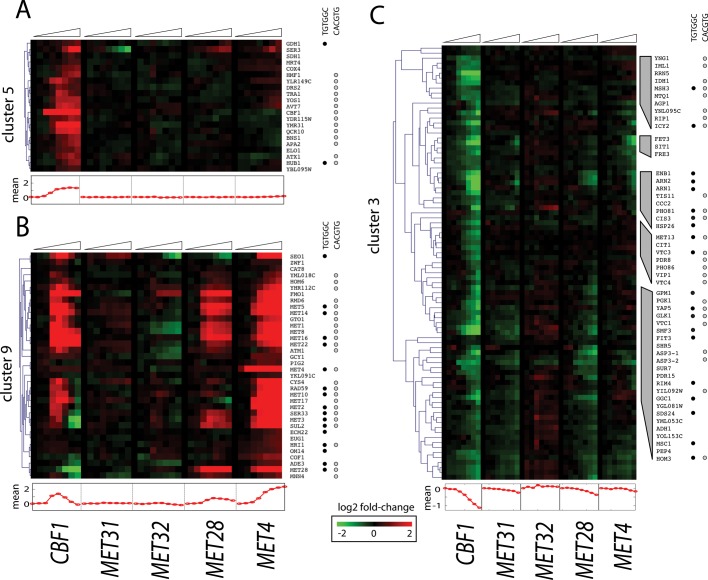
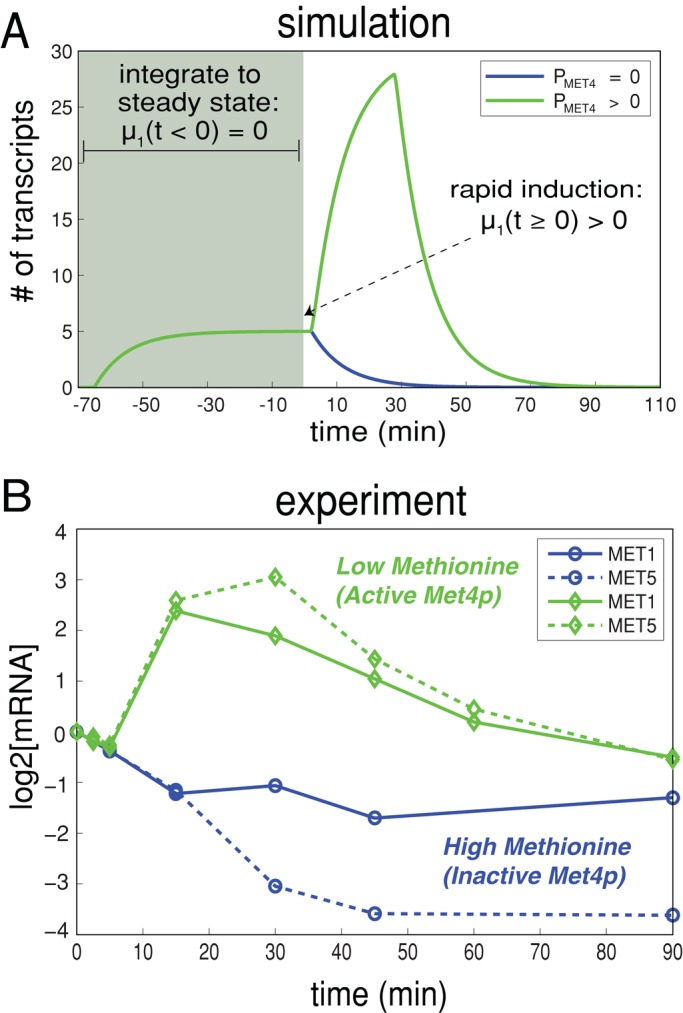
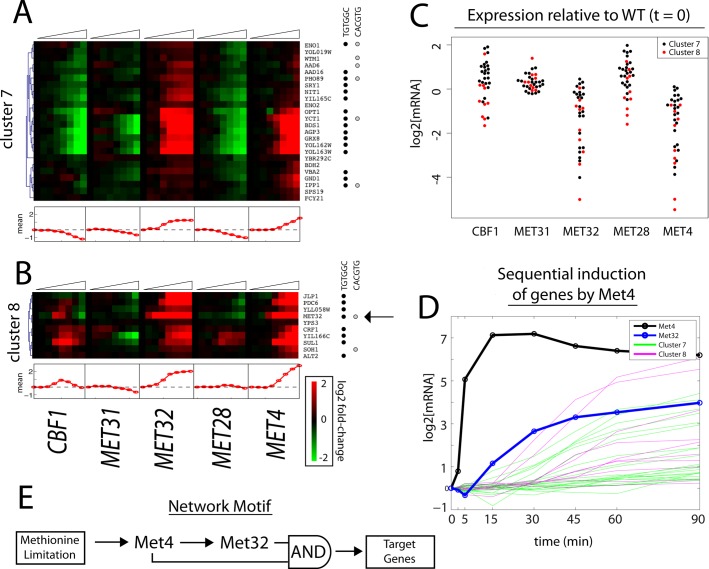
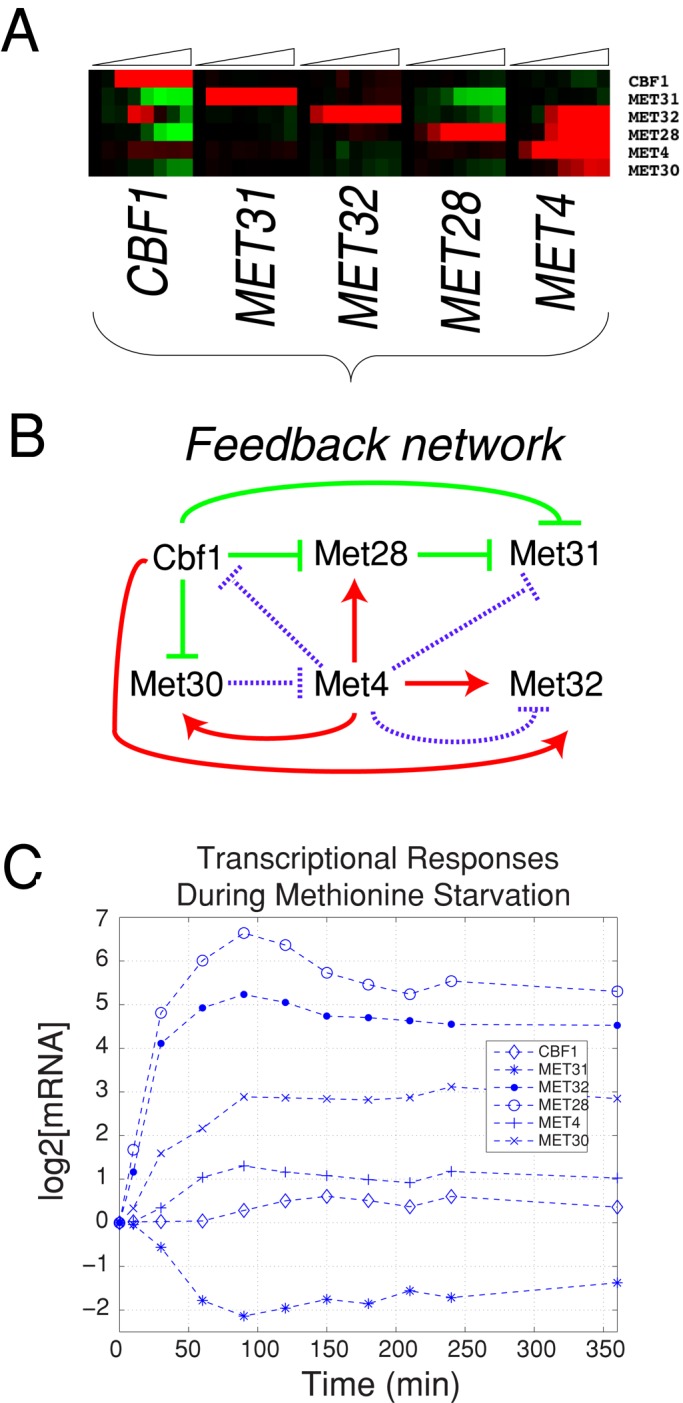
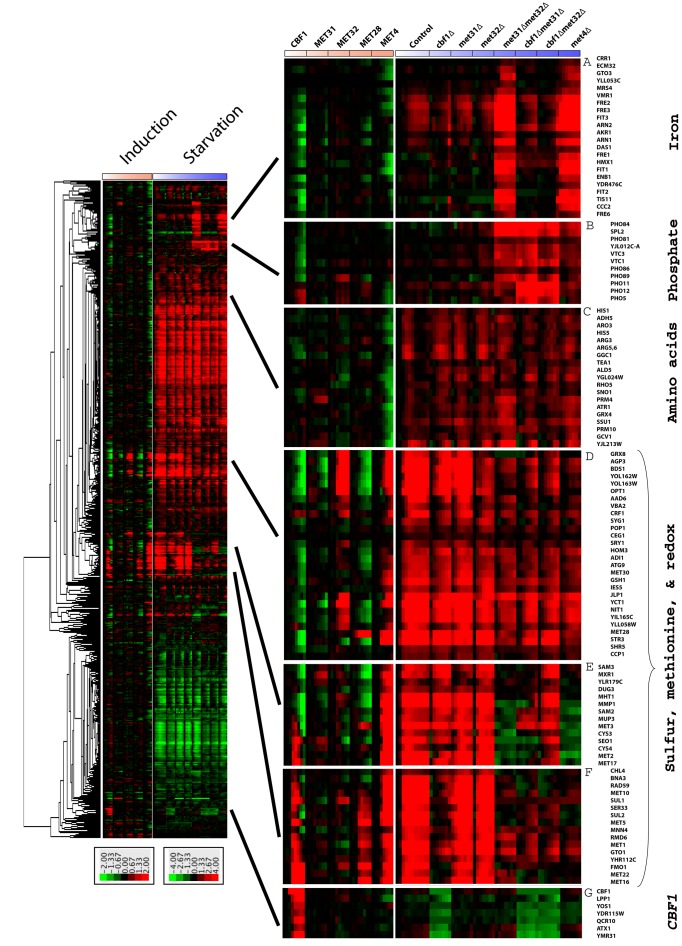
In yeast, the pathways of sulfur assimilation are combinatorially controlled by five transcriptional regulators (three DNA-binding proteins [Met31p, Met32p, and Cbf1p], an activator [Met4p], and a cofactor [Met28p]) and a ubiquitin ligase subunit (Met30p). This regulatory system exerts combinatorial control not only over sulfur assimilation and methionine biosynthesis, but also on many other physiological functions in the cell. Recently we characterized a gene induction system that, upon the addition of an inducer, results in near-immediate transcription of a gene of interest under physiological conditions. We used this to perturb levels of single transcription factors during steady-state growth in chemostats, which facilitated distinction of direct from indirect effects of individual factors dynamically through quantification of the subsequent changes in genome-wide patterns of gene expression. We were able to show directly that Cbf1p acts sometimes as a repressor and sometimes as an activator. We also found circumstances in which Met31p/Met32p function as repressors, as well as those in which they function as activators. We elucidated and numerically modeled feedback relationships among the regulators, notably feedforward regulation of Met32p (but not Met31p) by Met4p that generates dynamic differences in abundance that can account for the differences in function of these two proteins despite their identical binding sites.
在酵母中,硫同化途径通过五种转录调节剂(三个 DNA 结合蛋白[Met31p、Met32p 和 Cbf1p]、一个激活剂[Met4p]和一个辅因子[Met28p])和一个泛素连接酶亚基(Met30p)进行组合控制。该调节系统不仅对硫同化和蛋氨酸生物合成进行组合控制,而且对细胞中的许多其他生理功能进行组合控制。最近,我们描述了一种基因诱导系统,该系统在添加诱导剂后,在生理条件下几乎立即转录感兴趣的基因。我们使用该系统在恒化器中的稳态生长过程中扰动单个转录因子的水平,这有助于通过定量随后基因组范围内基因表达模式的变化来区分单个因子的直接和间接影响。我们能够直接表明 Cbf1p 有时作为抑制剂,有时作为激活剂。我们还发现了 Met31p/Met32p 作为抑制剂起作用的情况,以及它们作为激活剂起作用的情况。我们阐明并数值建模了调节剂之间的反馈关系,特别是 Met4p 对 Met32p(而不是 Met31p)的前馈调节,这种调节产生了丰度的动态差异,这些差异可以解释这两种蛋白质尽管具有相同的结合位点但功能不同的原因。