Department of Biology, The University of North Carolina at Chapel Hill, Chapel Hill, North Carolina, United States of America.
PLoS Genet. 2012 Jun;8(6):e1002756. doi: 10.1371/journal.pgen.1002756. Epub 2012 Jun 28.
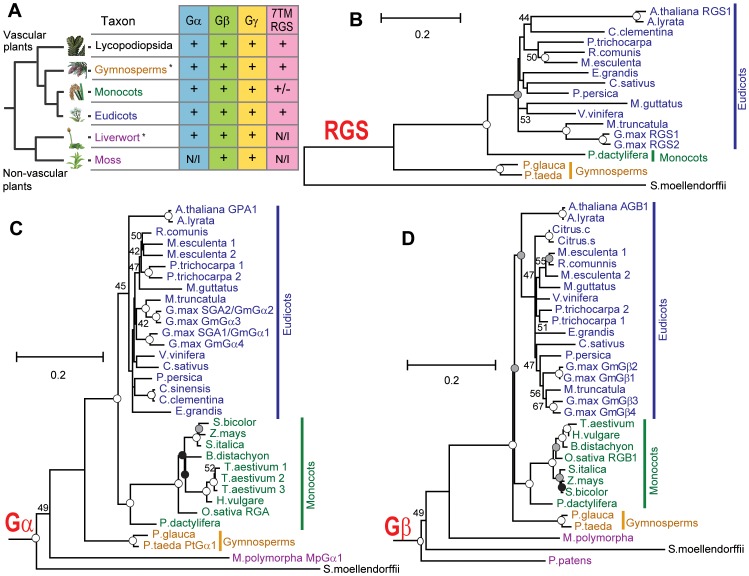
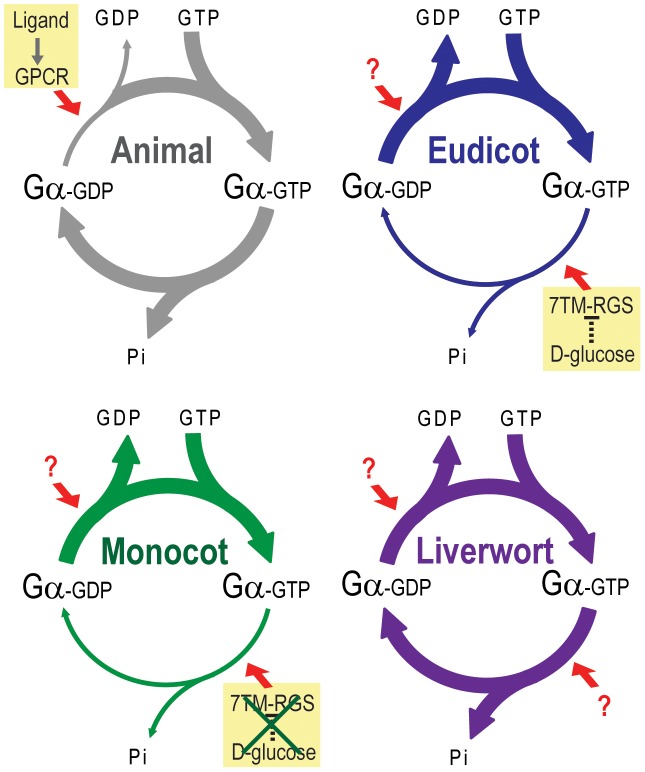
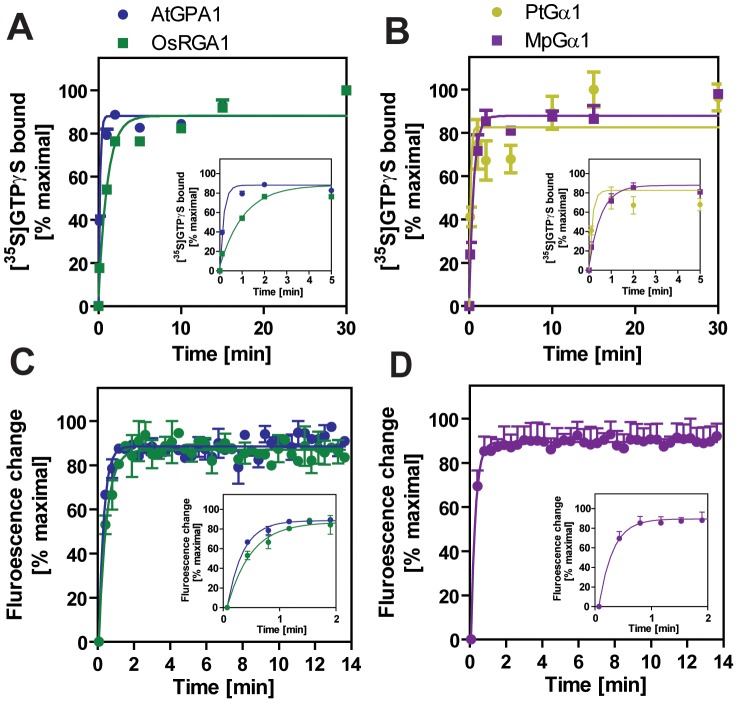
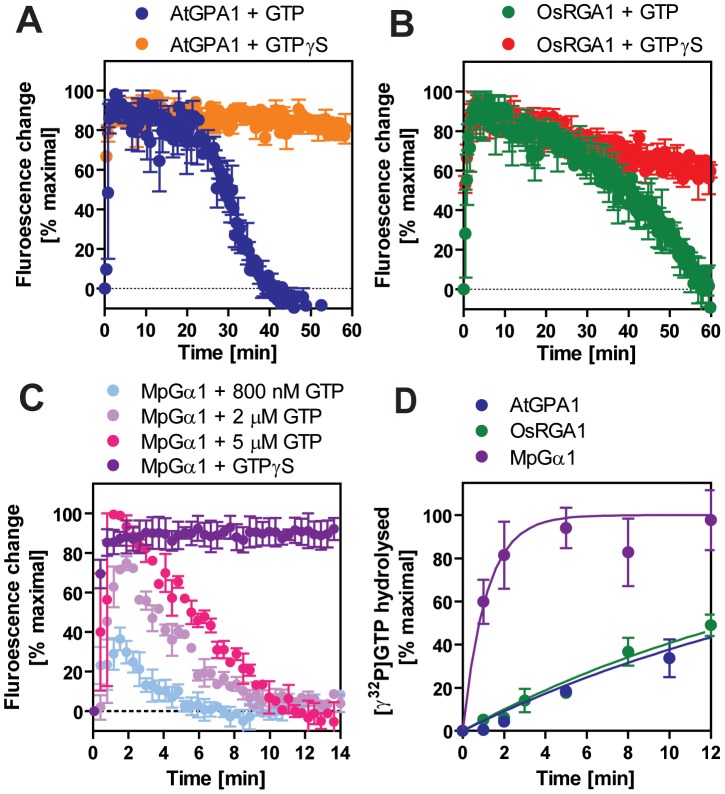
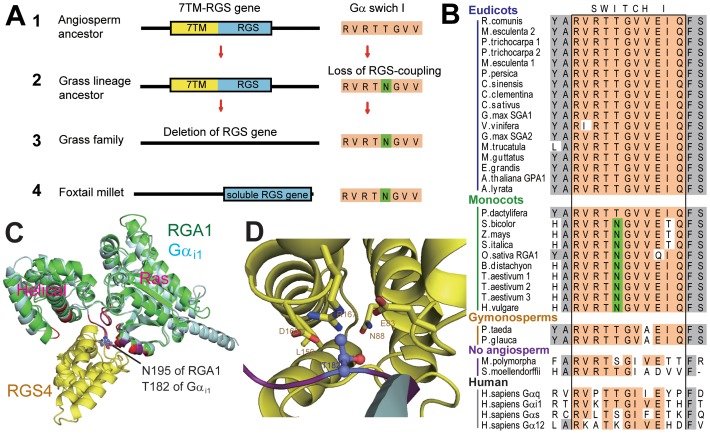
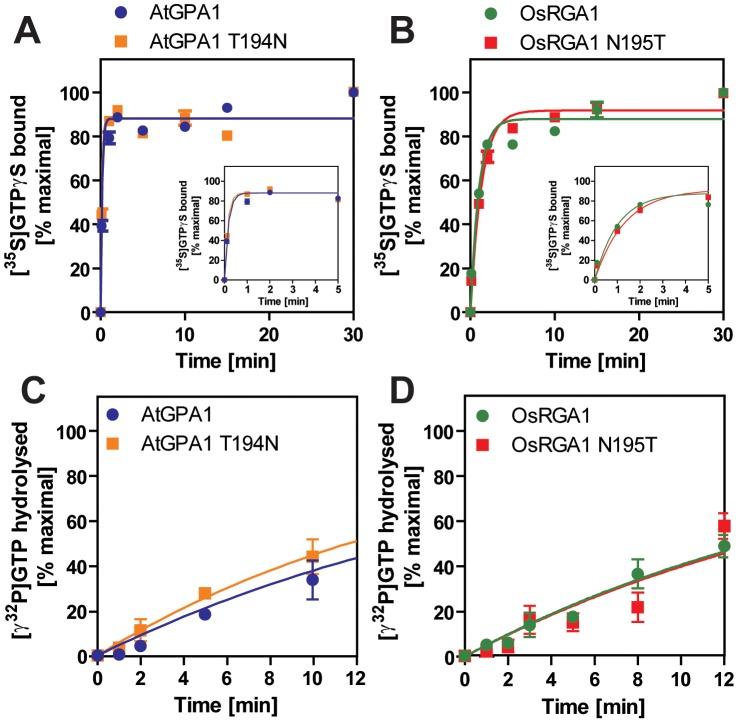
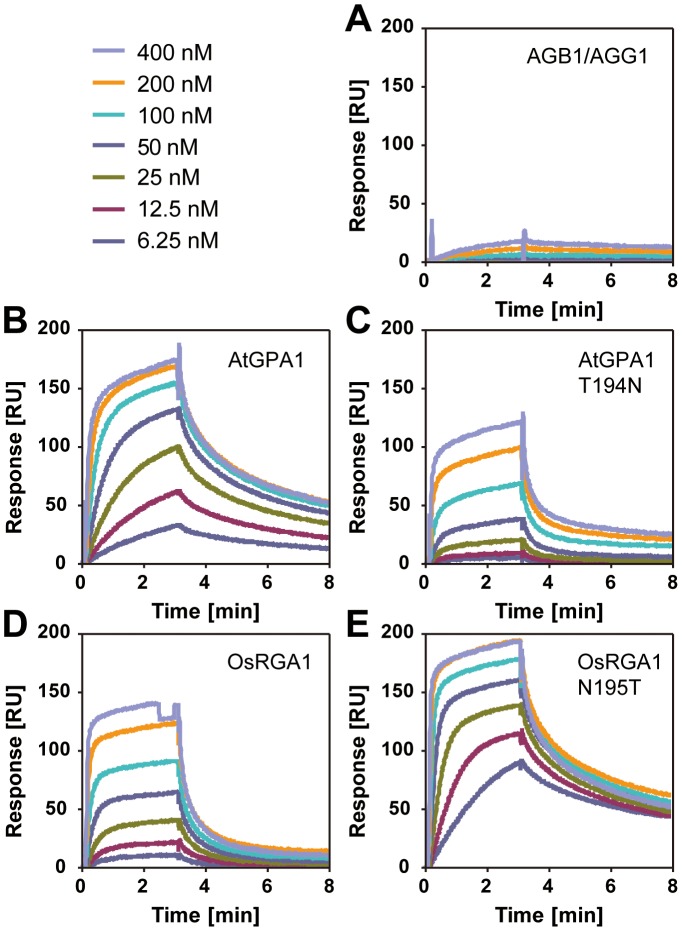
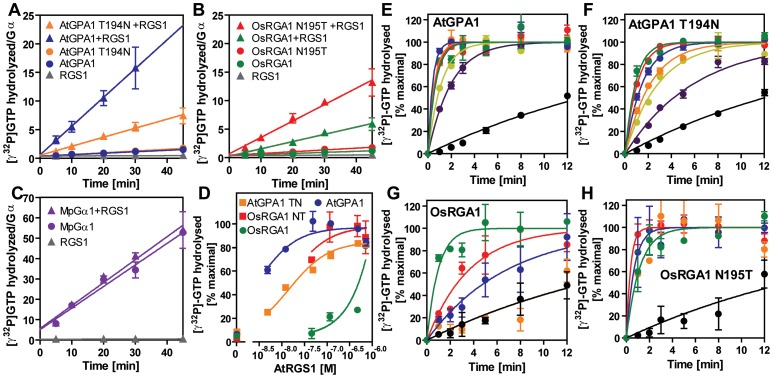
Animal heterotrimeric G proteins are activated by guanine nucleotide exchange factors (GEF), typically seven transmembrane receptors that trigger GDP release and subsequent GTP binding. In contrast, the Arabidopsis thaliana G protein (AtGPA1) rapidly activates itself without a GEF and is instead regulated by a seven transmembrane Regulator of G protein Signaling (7TM-RGS) protein that promotes GTP hydrolysis to reset the inactive (GDP-bound) state. It is not known if this unusual activation is a major and constraining part of the evolutionary history of G signaling in eukaryotes. In particular, it is not known if this is an ancestral form or if this mechanism is maintained, and therefore constrained, within the plant kingdom. To determine if this mode of signal regulation is conserved throughout the plant kingdom, we analyzed available plant genomes for G protein signaling components, and we purified individually the plant components encoded in an informative set of plant genomes in order to determine their activation properties in vitro. While the subunits of the heterotrimeric G protein complex are encoded in vascular plant genomes, the 7TM-RGS genes were lost in all investigated grasses. Despite the absence of a Gα-inactivating protein in grasses, all vascular plant Gα proteins examined rapidly released GDP without a receptor and slowly hydrolyzed GTP, indicating that these Gα are self-activating. We showed further that a single amino acid substitution found naturally in grass Gα proteins reduced the Gα-RGS interaction, and this amino acid substitution occurred before the loss of the RGS gene in the grass lineage. Like grasses, non-vascular plants also appear to lack RGS proteins. However, unlike grasses, one representative non-vascular plant Gα showed rapid GTP hydrolysis, likely compensating for the loss of the RGS gene. Our findings, the loss of a regulatory gene and the retention of the "self-activating" trait, indicate the existence of divergent Gα regulatory mechanisms in the plant kingdom. In the grasses, purifying selection on the regulatory gene was lost after the physical decoupling of the RGS protein and its cognate Gα partner. More broadly these findings show extreme divergence in Gα activation and regulation that played a critical role in the evolution of G protein signaling pathways.
动物异三聚体 G 蛋白被鸟嘌呤核苷酸交换因子 (GEF) 激活,通常是七次跨膜受体,可触发 GDP 释放和随后的 GTP 结合。相比之下,拟南芥 G 蛋白 (AtGPA1) 无需 GEF 即可快速自身激活,而是受七次跨膜 G 蛋白信号转导调节因子 (7TM-RGS) 蛋白调节,该蛋白促进 GTP 水解以重置非活性 (GDP 结合) 状态。目前尚不清楚这种不寻常的激活是否是真核生物 G 信号转导进化历史的主要和约束部分。特别是,目前尚不清楚这是一种原始形式,还是这种机制在植物界中得到维持和因此受到限制。为了确定这种信号调节模式是否在整个植物界中保守,我们分析了可用的植物基因组中的 G 蛋白信号转导成分,并单独纯化了在一组信息丰富的植物基因组中编码的植物成分,以便在体外确定它们的激活特性。虽然异三聚体 G 蛋白复合物的亚基在维管植物基因组中被编码,但在所有研究的禾本科植物中,7TM-RGS 基因都丢失了。尽管在禾本科植物中没有 Gα 失活蛋白,但所有被检查的维管植物 Gα 蛋白在没有受体的情况下迅速释放 GDP,并缓慢水解 GTP,这表明这些 Gα 是自我激活的。我们进一步表明,在禾本科植物 Gα 蛋白中自然存在的单个氨基酸取代降低了 Gα-RGS 相互作用,并且该氨基酸取代发生在禾本科植物谱系中 RGS 基因丢失之前。与禾本科植物一样,非维管束植物似乎也缺乏 RGS 蛋白。然而,与禾本科植物不同的是,一种代表性的非维管束植物 Gα 表现出快速的 GTP 水解,可能补偿了 RGS 基因的缺失。我们的发现,即调节基因的丢失和“自我激活”特性的保留,表明在植物界中存在不同的 Gα 调节机制。在禾本科植物中,RGS 蛋白与其同源 Gα 伴侣的物理分离后,对调节基因的纯化选择丢失。更广泛地说,这些发现表明 Gα 激活和调节的极端分歧在 G 蛋白信号转导途径的进化中发挥了关键作用。