Department of Neuroscience and Physiology, SUNY Upstate Medical University, 750 E. Adams St, Syracuse, NY 13210, USA.
Neural Dev. 2012 Jul 7;7:25. doi: 10.1186/1749-8104-7-25.
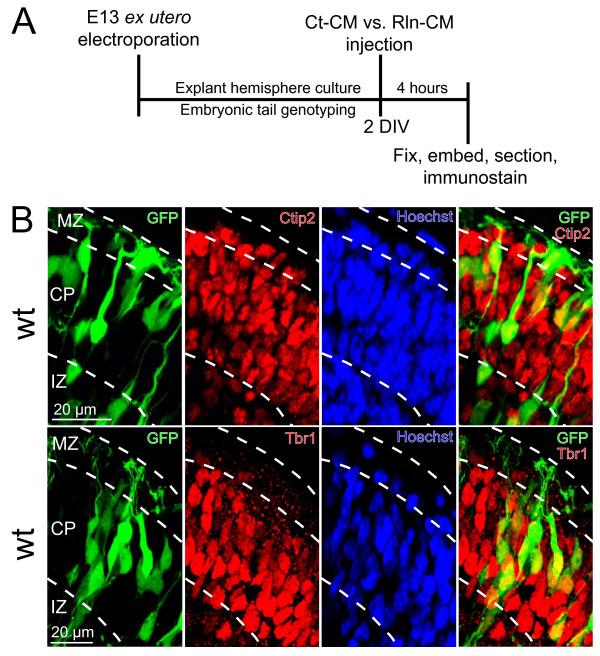
The secreted ligand Reelin is believed to regulate the translocation of prospective layer 6 (L6) neocortical neurons into the preplate, a loose layer of pioneer neurons that overlies the ventricular zone. Recent studies have also suggested that Reelin controls neuronal orientation and polarized dendritic growth during this period of early cortical development. To explicitly characterize and quantify how Reelin controls this critical aspect of neurite initiation and growth we used a new ex utero explant model of early cortical development to selectively label a subset of L6 cortical neurons for complete 3-D reconstruction.
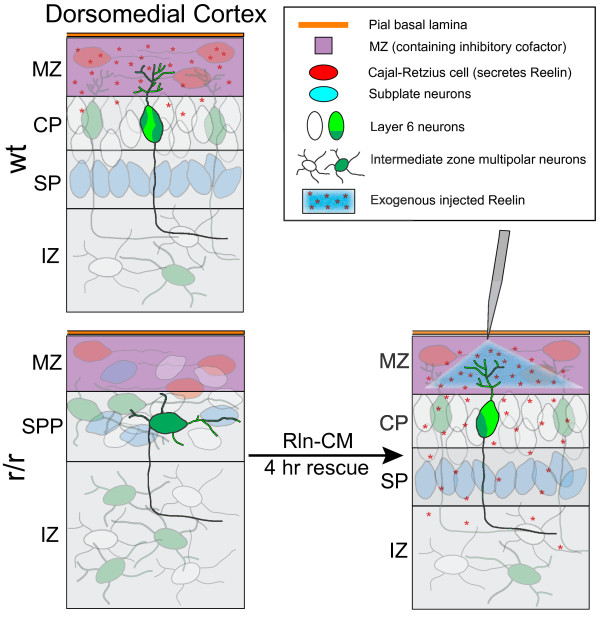
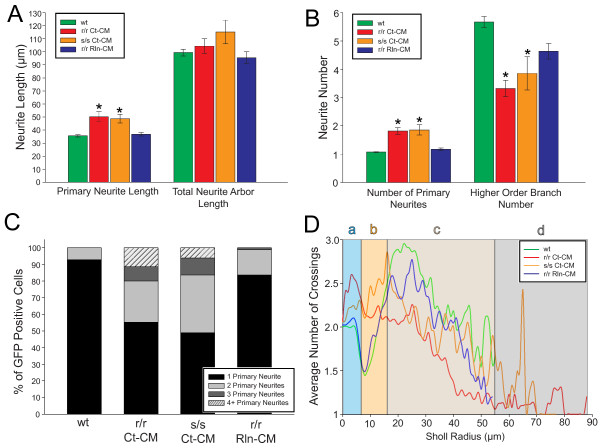
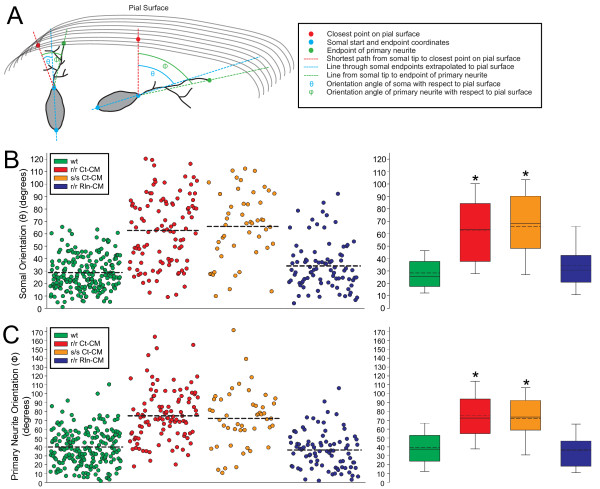
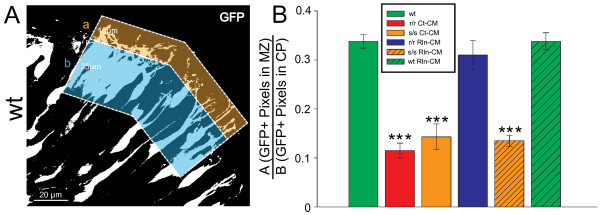
The total neurite arbor sizes of neurons in Reelin-deficient (reeler mutant) and Dab1-deficient (Reelin-non-responsive scrambler mutant) cortices were quantified and unexpectedly were not different than control arbor lengths (p = 0.51). For each mutant, however, arbor organization was markedly different: mutant neurons manifested more primary processes (neurites emitted directly from the soma) than wild type, and these neurites were longer and displayed less branching. Reeler and scrambler mutant neurites extended tangentially rather than radially, and the Golgi apparatus that normally invests the apical neurite was compact in both reeler and scrambler mutants. Mutant cortices also exhibited a neurite "exclusion zone" which was relatively devoid of L6 neuron neurites and extended at least 15 μm beneath the pial surface, an area corresponding to the marginal zone (MZ) in the wild type explants. The presence of an exclusion zone was also indicated in the orientation of mutant primary neurite and neuronal somata, which failed to adopt angles within ~20˚ of the radial line to the pial surface. Injection of recombinant Reelin to reeler, but not scrambler, mutant cortices fully rescued soma orientation, Golgi organization, and dendritic projection defects within four hrs.
These findings indicate Reelin promotes directional dendritic growth into the MZ, an otherwise exclusionary zone for L6 neurites.
分泌配体 Reelin 被认为可以调节前皮质层(preplate)中新生的 6 层(L6)皮质神经元的迁移,前皮质层是位于脑室区之上的松散的先驱神经元层。最近的研究还表明,Reelin 在这段早期皮质发育过程中控制神经元的定向和极化树突生长。为了明确描述和量化 Reelin 如何控制神经突起始和生长的这一关键方面,我们使用了一种新的离体皮质外植体模型来选择性标记一小部分 L6 皮质神经元,以进行完整的 3-D 重建。
Reelin 缺失(reeler 突变体)和 Dab1 缺失(Reelin 非反应性 scrambler 突变体)皮质中的神经元的总神经突树突大小进行了量化,出乎意料的是,与对照树突长度没有差异(p=0.51)。然而,对于每种突变体,树突组织都明显不同:突变体神经元比野生型表现出更多的初级突起(直接从神经元体发出的神经突),这些神经突更长,分支更少。reeler 和 scrambler 突变体的神经突呈切线延伸,而不是放射状延伸,正常覆盖顶突的高尔基器在 reeler 和 scrambler 突变体中都很紧凑。突变体皮质还表现出神经突“排斥区”,该区域相对缺乏 L6 神经元的神经突,并且至少在软脑膜表面下延伸 15μm,该区域与野生型外植体的边缘区(MZ)相对应。排斥区的存在也表现在突变体初级神经突和神经元体的方向上,它们未能在与软脑膜表面成~20˚的角度内形成角度。将重组 Reelin 注射到 reeler 但不是 scrambler 突变体皮质中,在 4 小时内完全挽救了体定向、高尔基组织和树突投射缺陷。
这些发现表明 Reelin 促进定向树突生长进入 MZ,这是 L6 神经突的一个排斥区。