Division of Biochemistry, The Netherlands Cancer Institute, Amsterdam, The Netherlands.
EMBO J. 2012 Oct 3;31(19):3833-44. doi: 10.1038/emboj.2012.217. Epub 2012 Aug 3.
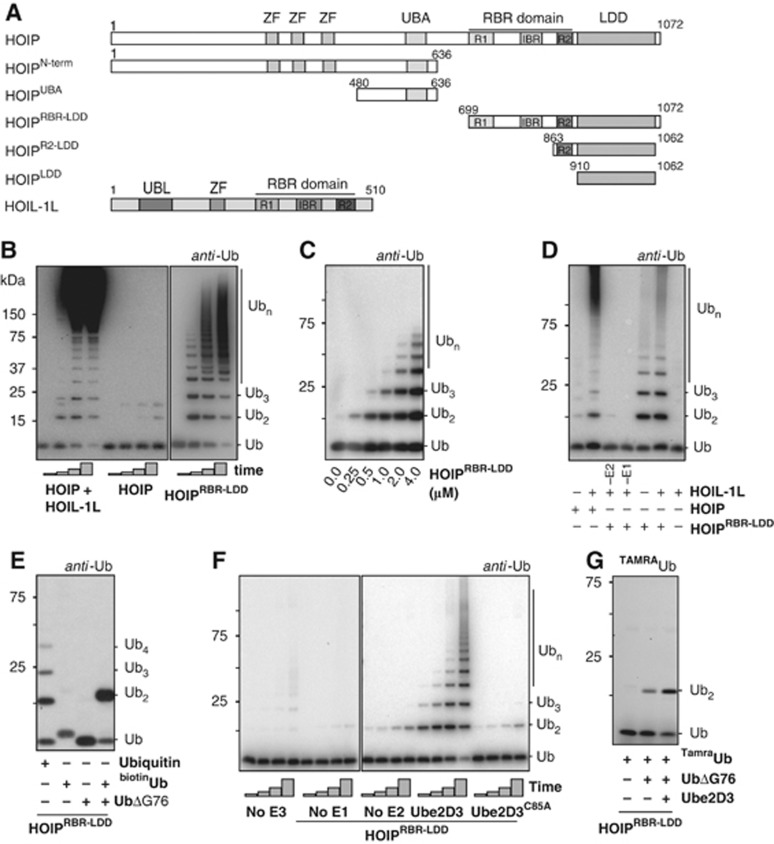
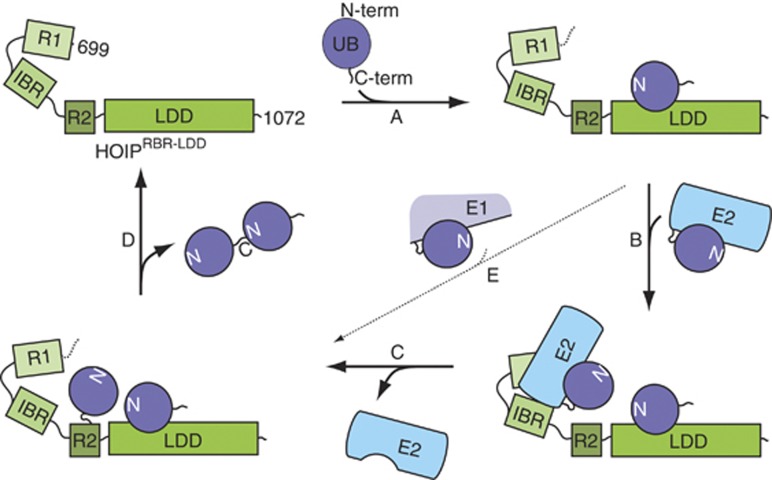
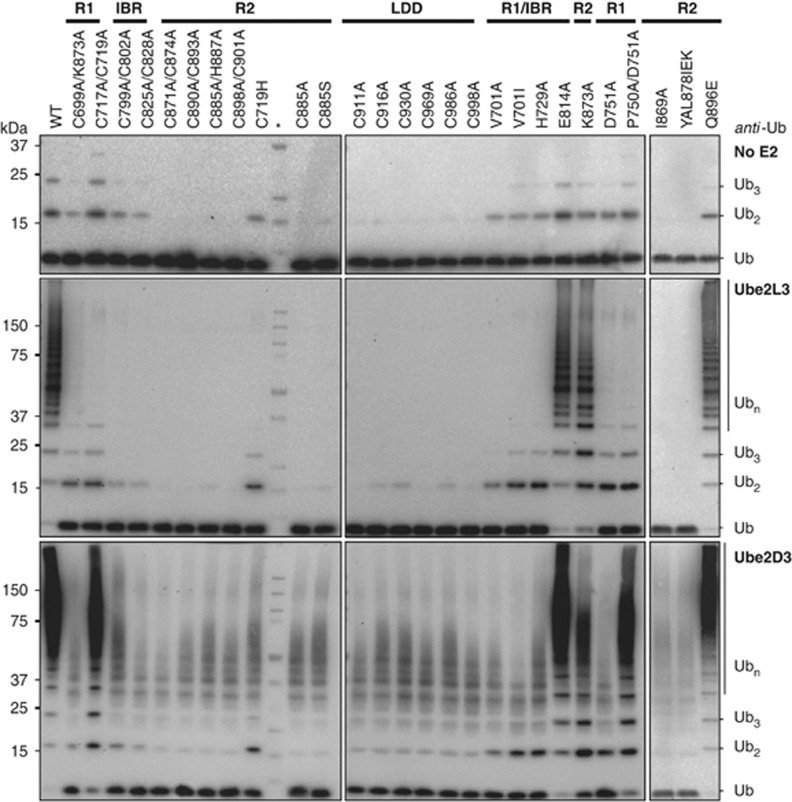
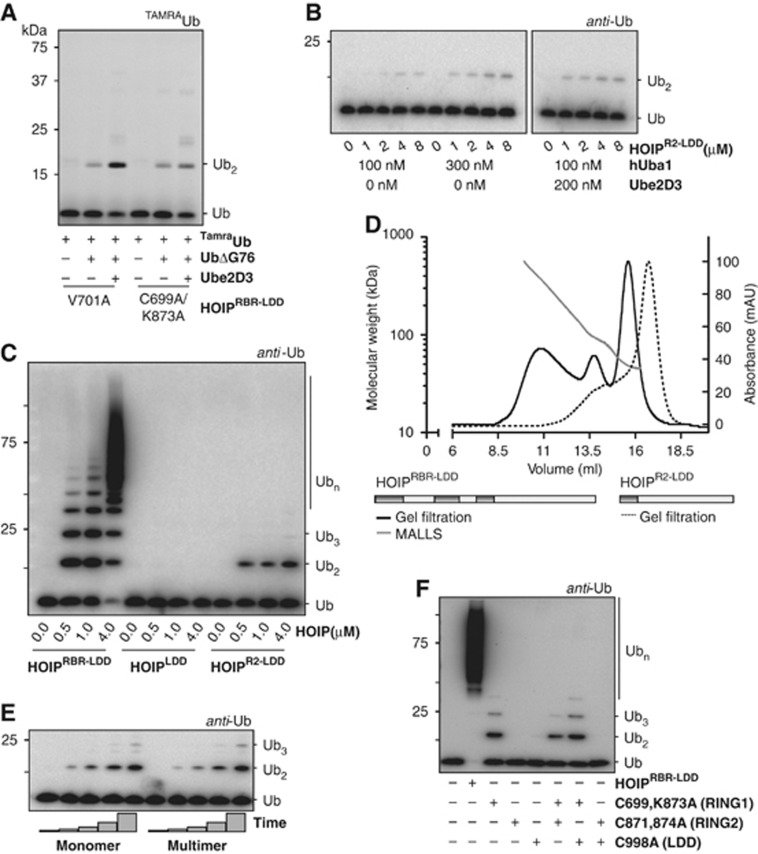
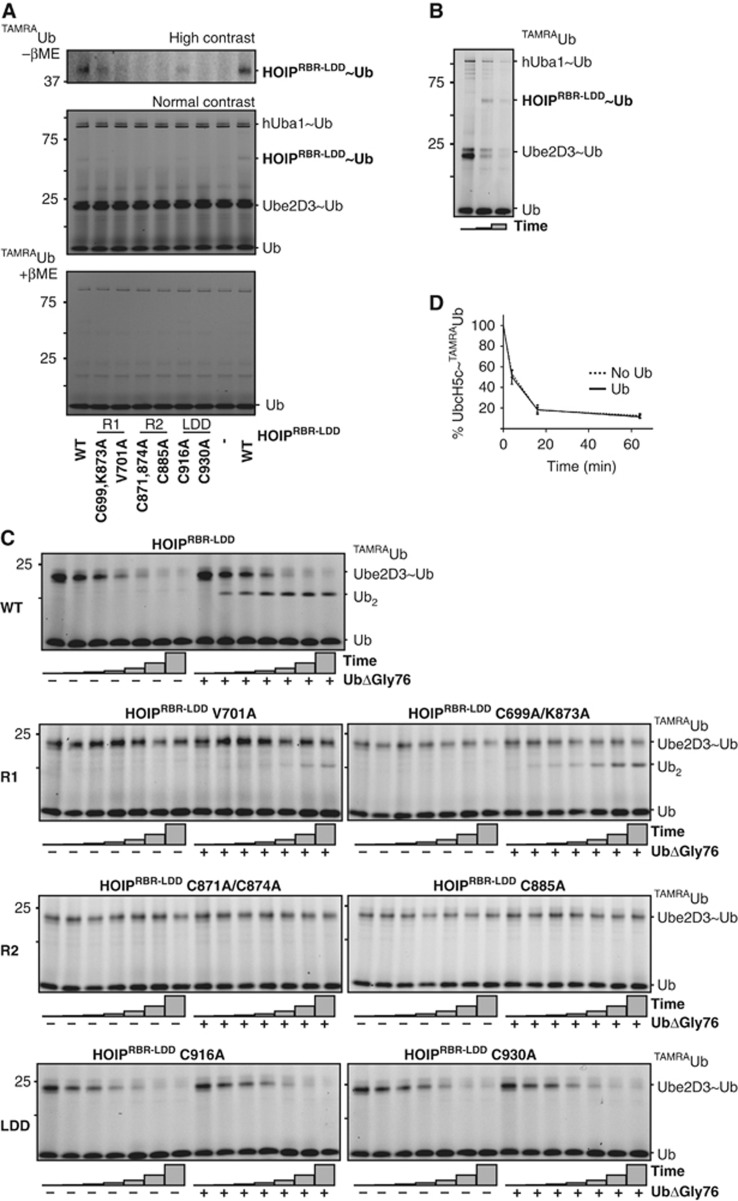
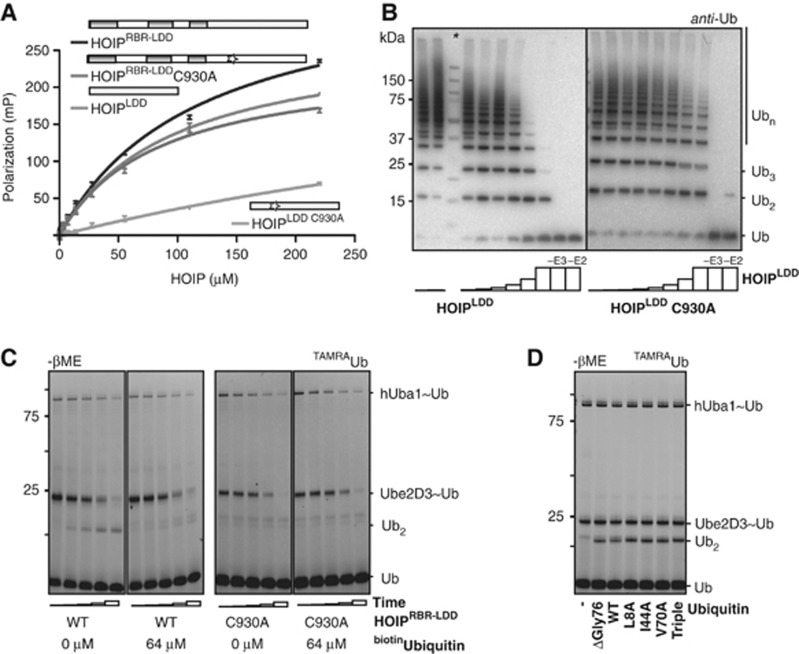
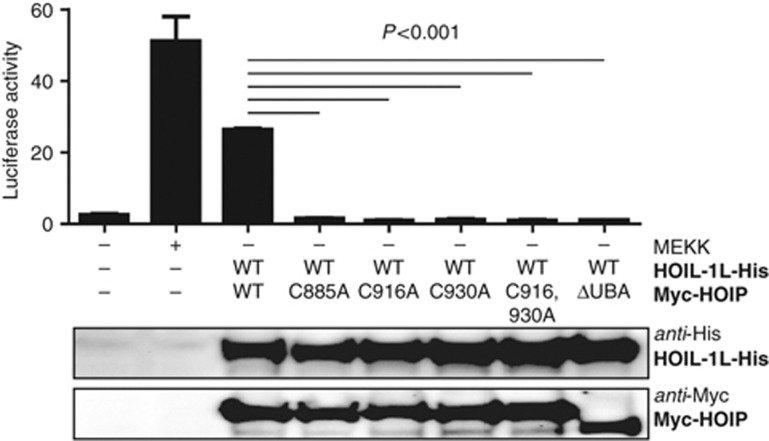
Activation of the NF-κB pathway requires the formation of Met1-linked 'linear' ubiquitin chains on NEMO, which is catalysed by the Linear Ubiquitin Chain Assembly Complex (LUBAC) E3 consisting of HOIP, HOIL-1L and Sharpin. Here, we show that both LUBAC catalytic activity and LUBAC specificity for linear ubiquitin chain formation are embedded within the RING-IBR-RING (RBR) ubiquitin ligase subunit HOIP. Linear ubiquitin chain formation by HOIP proceeds via a two-step mechanism involving both RING and HECT E3-type activities. RING1-IBR catalyses the transfer of ubiquitin from the E2 onto RING2, to transiently form a HECT-like covalent thioester intermediate. Next, the ubiquitin is transferred from HOIP onto the N-terminus of a target ubiquitin. This transfer is facilitated by a unique region in the C-terminus of HOIP that we termed 'Linear ubiquitin chain Determining Domain' (LDD), which may coordinate the acceptor ubiquitin. Consistent with this mechanism, the RING2-LDD region was found to be important for NF-κB activation in cellular assays. These data show how HOIP combines a general RBR ubiquitin ligase mechanism with unique, LDD-dependent specificity for producing linear ubiquitin chains.
NF-κB 通路的激活需要 NEMO 上形成 Met1 连接的“线性”泛素链,这是由包含 HOIP、HOIL-1L 和 Sharpin 的线性泛素链组装复合物(LUBAC)E3 催化的。在这里,我们表明,LUBAC 的催化活性和线性泛素链形成的特异性都嵌入在 RING-IBR-RING(RBR)泛素连接酶亚基 HOIP 中。HOIP 的线性泛素链形成通过涉及 RING 和 HECT E3 型活性的两步机制进行。RING1-IBR 催化将泛素从 E2 转移到 RING2 上,以短暂形成 HECT 样共价硫酯中间物。接下来,泛素从 HOIP 转移到靶泛素的 N 端。这种转移由 HOIP 羧基末端的一个独特区域(我们称为“线性泛素链决定结构域”(LDD))促进,该区域可能协调受体泛素。与该机制一致,发现 RING2-LDD 区域对于细胞测定中的 NF-κB 激活很重要。这些数据显示了 HOIP 如何将通用的 RBR 泛素连接酶机制与独特的、依赖 LDD 的特异性结合起来,产生线性泛素链。