Department of Chemistry, University of California, Berkeley, CA 94720, USA.
Biochemistry. 2012 Sep 25;51(38):7488-95. doi: 10.1021/bi300456f. Epub 2012 Sep 12.
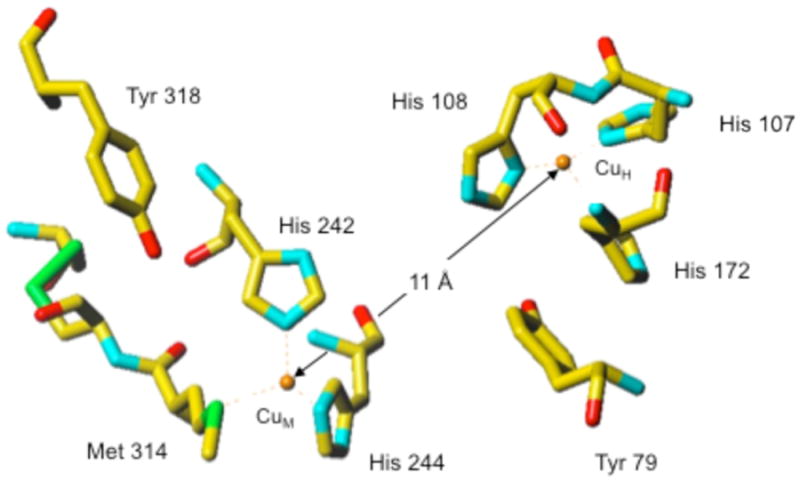
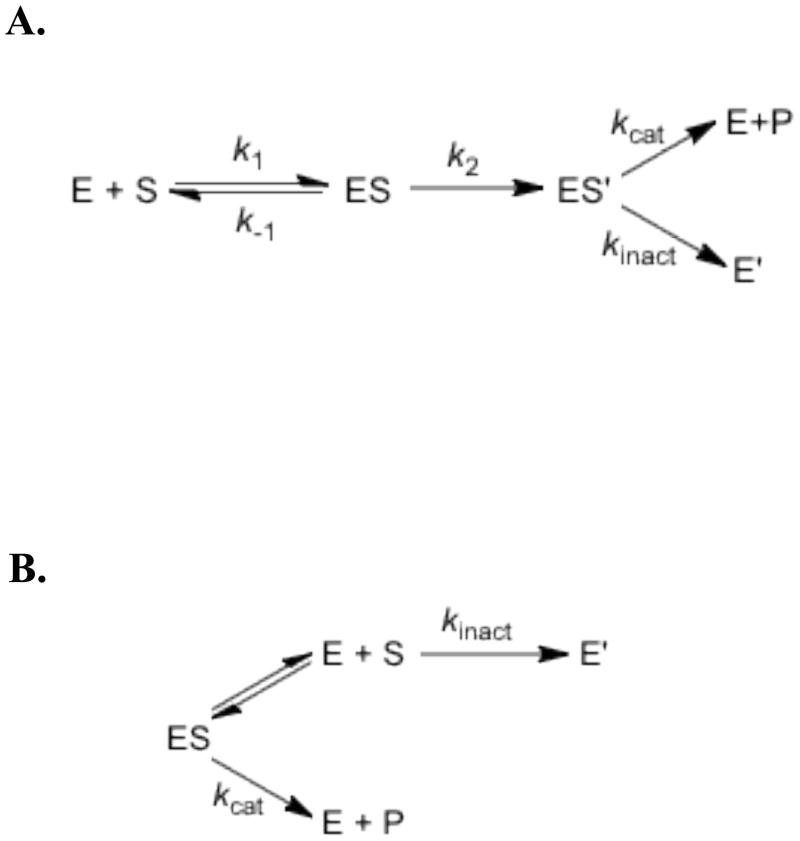
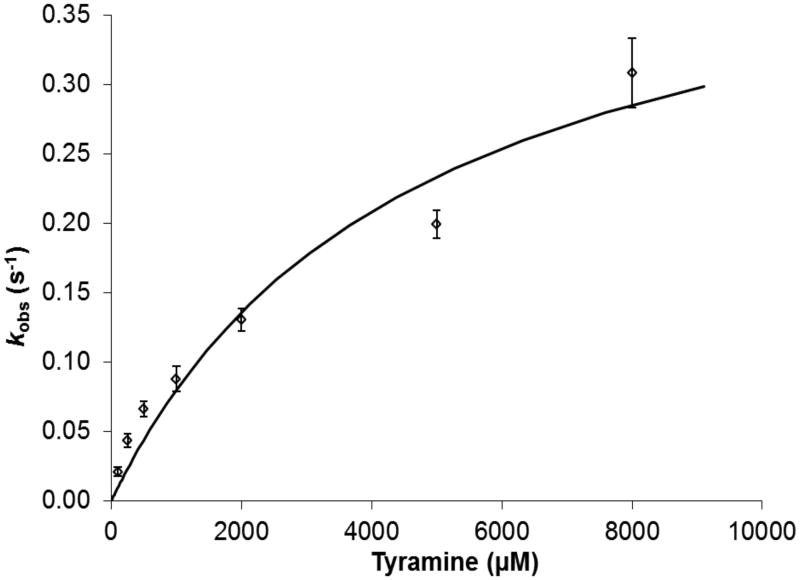
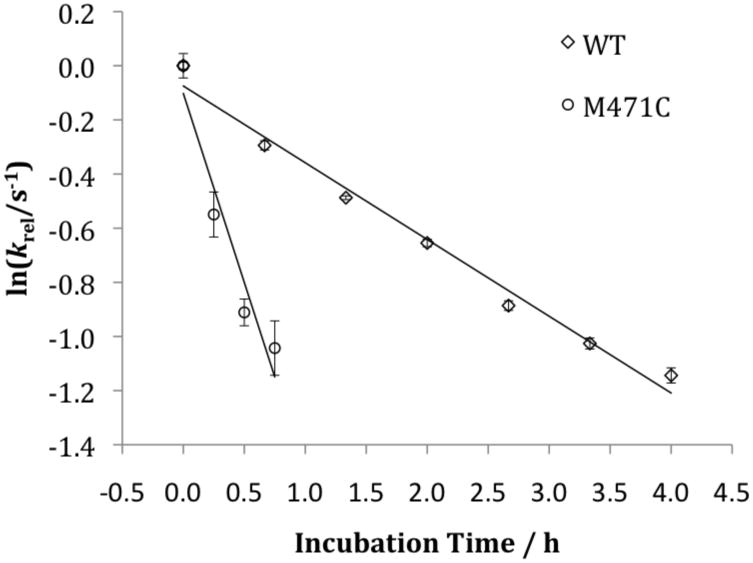
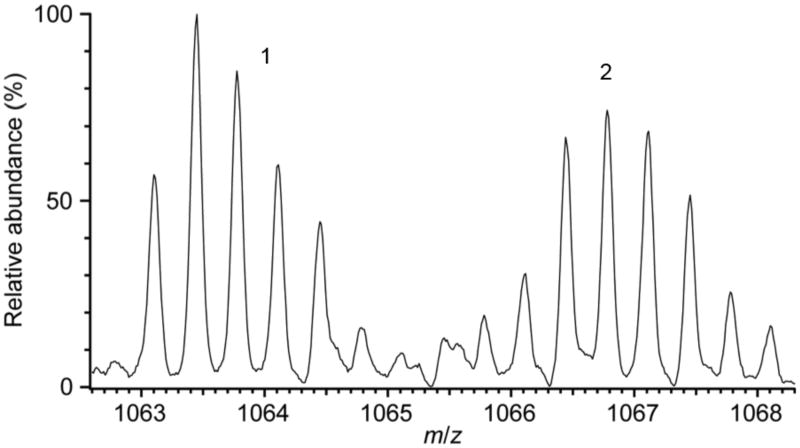
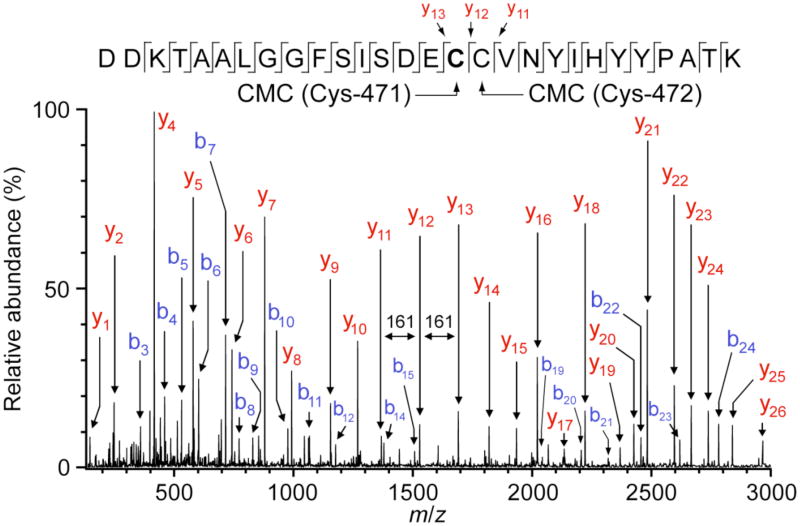
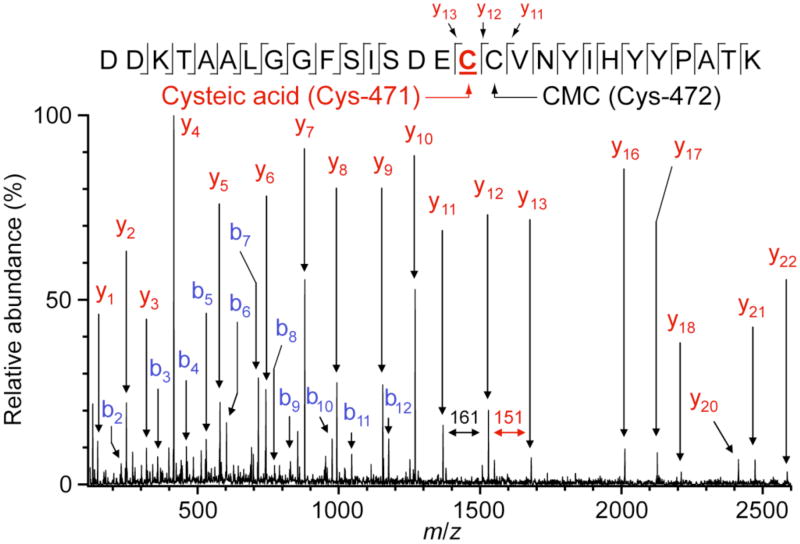
Tyramine β-monooxygenase (TβM), the insect homologue of dopamine β-monooxygenase, is a neuroregulatory enzyme that catalyzes the β-hydroxylation of tyramine to yield octopamine. Mutation of the methionine (Met) ligand to Cu(M) of TβM, Met471Cys, yielded a form of TβM that is catalytically active but susceptible to inactivation during turnover [Hess, C. R., Wu, Z., Ng, A., Gray, E. E., McGuirl, M. M., and Klinman, J. P. (2008) J. Am. Chem. Soc. 130, 11939-11944]. Further, although the wild-type (WT) enzyme undergoes coordination of Met471 to Cu(M) in its reduced form, the generation of Met471Cys almost completely eliminates this interaction [Hess, C. R., Klinman, J. P., and Blackburn, N. J. (2010) J. Biol. Inorg. Chem. 15, 1195-1207]. The aim of this study is to identify the chemical consequence of the poor ability of Cys to coordinate Cu(M). We show that Met471Cys TβM is ~5-fold more susceptible to inactivation than the WT enzyme in the presence of the cosubstrate/reductant ascorbate and that this process is not facilitated by the substrate tyramine. The resulting 50-fold smaller ratio for turnover to inactivation in the case of Met471Cys prevents full turnover of the substrate under all conditions examined. Liquid chromatography-tandem mass spectrometry analysis of proteolytic digests of inactivated Met471Cys TβM leads to the identification of cysteic acid at position 471. While both Met and Cys side chains are expected to be similarly subject to oxidative damage in proteins, the enhanced reactivity of Met471Cys toward solution oxidants in TβM is attributed to its weaker interaction with Cu(I)(M).
酪胺 β-单加氧酶(TβM)是多巴胺 β-单加氧酶的昆虫同源物,是一种神经调节酶,可催化酪胺的β-羟化生成章鱼胺。TβM 的蛋氨酸(Met)配体到 Cu(M)的突变,Met471Cys,产生了一种具有催化活性但在周转过程中容易失活的 TβM 形式[Hess,C.R.,Wu,Z.,Ng,A.,Gray,E.E.,McGuirl,M.M.,和 Klinman,J.P.(2008)J.Am.Chem.Soc.130,11939-11944]。此外,尽管野生型(WT)酶在其还原形式下经历 Met471 与 Cu(M)的配位,但 Met471Cys 的产生几乎完全消除了这种相互作用[Hess,C.R.,Klinman,J.P.,和 Blackburn,N.J.(2010)J.Biol.Inorg.Chem.15,1195-1207]。本研究的目的是确定 Cys 配位 Cu(M)能力差的化学后果。我们表明,Met471Cys TβM 在存在辅助因子/还原剂抗坏血酸的情况下比 WT 酶更容易失活,并且该过程不受底物酪胺的促进。在 Met471Cys 的情况下,失活与周转的比率小 5 倍,可防止在所有检查的条件下完全转化底物。对失活的 Met471Cys TβM 的蛋白水解消化产物进行液相色谱-串联质谱分析,导致在位置 471 鉴定出半胱氨酸磺酸。虽然 Met 和 Cys 侧链都预计在蛋白质中受到类似的氧化损伤,但 TβM 中 Met471Cys 对溶液氧化剂的反应性增强归因于其与 Cu(I)(M)的相互作用较弱。