Department of Toxicogenetics, Leiden University Medical Center, Einthovenweg 20, 2333 ZC Leiden, The Netherlands.
Nature. 2012 Sep 27;489(7417):581-4. doi: 10.1038/nature11353. Epub 2012 Sep 9.
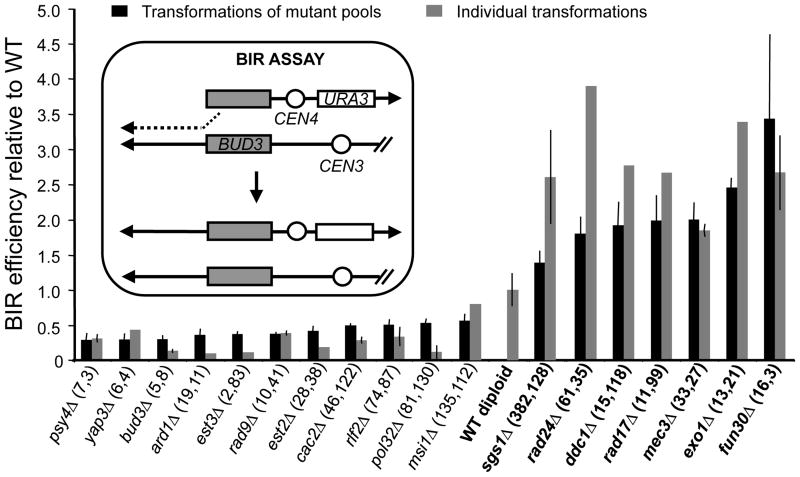
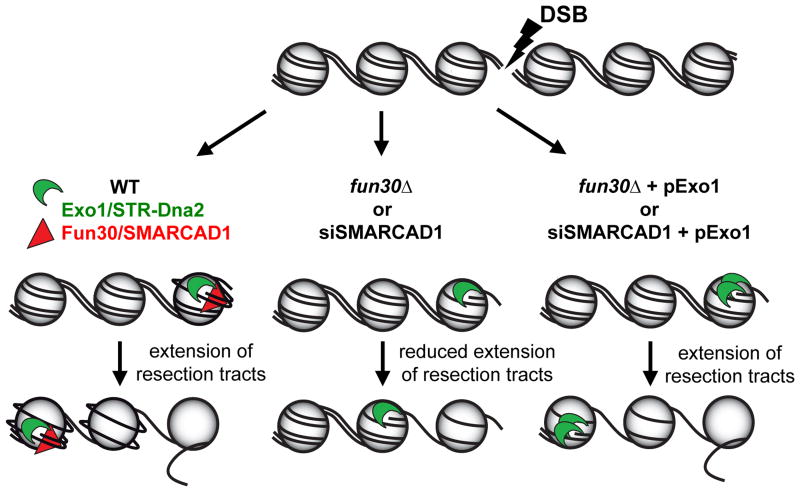
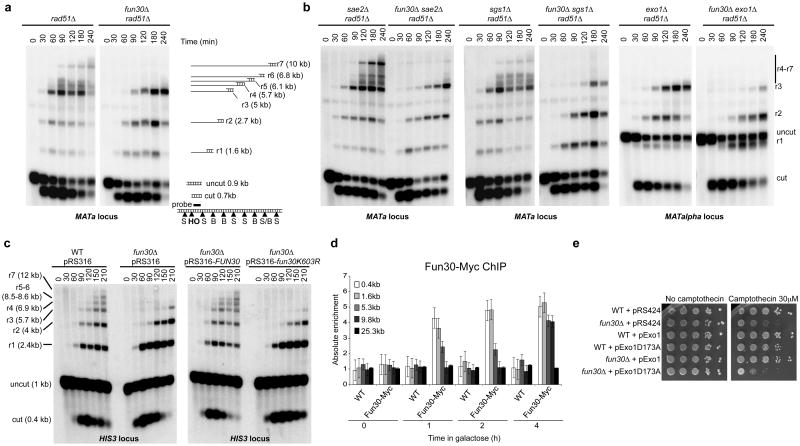
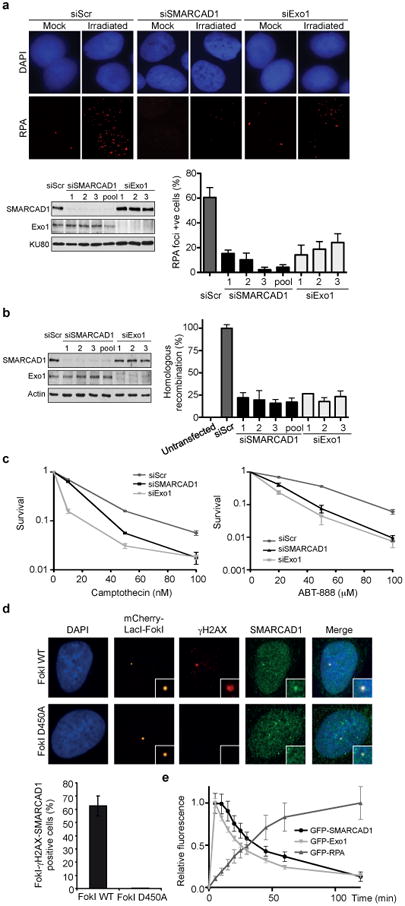
Several homology-dependent pathways can repair potentially lethal DNA double-strand breaks (DSBs). The first step common to all homologous recombination reactions is the 5'-3' degradation of DSB ends that yields the 3' single-stranded DNA required for the loading of checkpoint and recombination proteins. In yeast, the Mre11-Rad50-Xrs2 complex (Xrs2 is known as NBN or NBS1 in humans) and Sae2 (known as RBBP8 or CTIP in humans) initiate end resection, whereas long-range resection depends on the exonuclease Exo1, or the helicase-topoisomerase complex Sgs1-Top3-Rmi1 together with the endonuclease Dna2 (refs 1-6). DSBs occur in the context of chromatin, but how the resection machinery navigates through nucleosomal DNA is a process that is not well understood. Here we show that the yeast Saccharomyces cerevisiae Fun30 protein and its human counterpart SMARCAD1 (ref. 8), two poorly characterized ATP-dependent chromatin remodellers of the Snf2 ATPase family, are directly involved in the DSB response. Fun30 physically associates with DSB ends and directly promotes both Exo1- and Sgs1-dependent end resection through a mechanism involving its ATPase activity. The function of Fun30 in resection facilitates the repair of camptothecin-induced DNA lesions, although it becomes dispensable when Exo1 is ectopically overexpressed. Interestingly, SMARCAD1 is also recruited to DSBs, and the kinetics of recruitment is similar to that of EXO1. The loss of SMARCAD1 impairs end resection and recombinational DNA repair, and renders cells hypersensitive to DNA damage resulting from camptothecin or poly(ADP-ribose) polymerase inhibitor treatments. These findings unveil an evolutionarily conserved role for the Fun30 and SMARCAD1 chromatin remodellers in controlling end resection, homologous recombination and genome stability in the context of chromatin.
几种同源依赖性途径可以修复潜在致死性的 DNA 双链断裂 (DSB)。所有同源重组反应的第一步都是 5' - 3' 降解 DSB 末端,产生加载检查点和重组蛋白所需的 3' 单链 DNA。在酵母中,Mre11-Rad50-Xrs2 复合物(Xrs2 在人类中称为 NBN 或 NBS1)和 Sae2(在人类中称为 RBBP8 或 CTIP)启动末端切除,而长距离切除则依赖于外切核酸酶 Exo1,或解旋酶-拓扑异构酶复合物 Sgs1-Top3-Rmi1 与内切核酸酶 Dna2 一起(参考文献 1-6)。DSB 发生在染色质的背景下,但切除机制如何在核小体 DNA 中导航是一个尚未完全理解的过程。在这里,我们表明酵母酿酒酵母 Fun30 蛋白及其人类对应物 SMARCAD1(参考文献 8),这两种 Snf2 ATP 酶家族中功能尚未完全确定的 ATP 依赖性染色质重塑因子,直接参与 DSB 反应。Fun30 与 DSB 末端物理结合,并通过涉及其 ATP 酶活性的机制直接促进 Exo1 和 Sgs1 依赖性末端切除。Fun30 在切除中的功能促进了喜树碱诱导的 DNA 损伤的修复,尽管当 Exo1 异位过表达时,它变得可有可无。有趣的是,SMARCAD1 也被招募到 DSB 上,并且募集的动力学与 EXO1 相似。SMARCAD1 的缺失会损害末端切除和重组 DNA 修复,并使细胞对喜树碱或多聚 (ADP-核糖) 聚合酶抑制剂处理引起的 DNA 损伤变得敏感。这些发现揭示了 Fun30 和 SMARCAD1 染色质重塑因子在控制染色质背景下的末端切除、同源重组和基因组稳定性方面的进化保守作用。