Laboratory of Plasma Derivatives, Division of Hematology, Office of Blood Research and Review, Center for Biologics Evaluation and Research, Food and Drug Administration, FDA/CBER/OBRR/DH/LPD, HFM-345, 1401 Rockville Pike, Rockville, MD 20852, USA.
Virol J. 2012 Sep 24;9:217. doi: 10.1186/1743-422X-9-217.
A33 is a type II integral membrane protein expressed on the extracellular enveloped form of vaccinia virus (VACV). Passive transfer of A33-directed monoclonal antibodies or vaccination with an A33 subunit vaccine confers protection against lethal poxvirus challenge in animal models. Homologs of A33 are highly conserved among members of the Orthopoxvirus genus and are potential candidates for inclusion in vaccines or assays targeting extracellular enveloped virus activity. One monoclonal antibody directed against VACV A33, MAb-1G10, has been shown to target a conformation-dependent epitope. Interestingly, while it recognizes VACV A33 as well as the corresponding variola homolog, it does not bind to the monkeypox homolog. In this study, we utilized a random phage display library to investigate the epitope recognized by MAb-1G10 that is critical for facilitating cell-to-cell spread of the vaccinia virus.
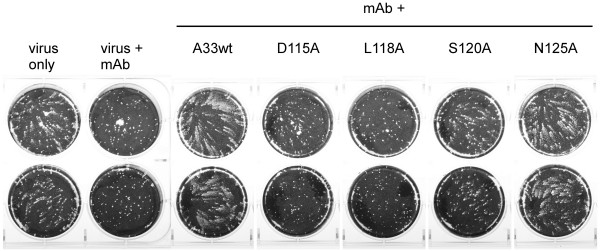
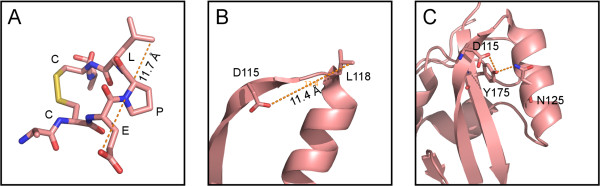
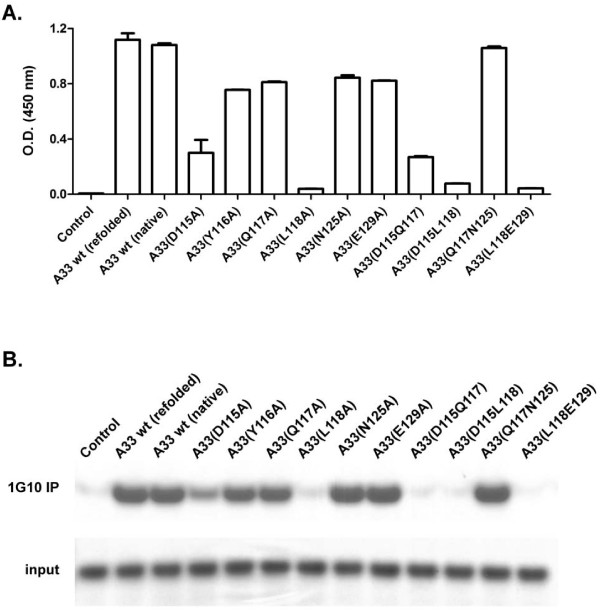
By screening with linear or conformational random phage libraries, we found that phages binding to MAb-1G10 display the consensus motif CEPLC, with a disulfide bond formed between two cysteine residues required for MAb-1G10 binding. Although the phage motif contained no linear sequences homologous to VACV A33, structure modeling and analysis suggested that residue D115 is important to form the minimal epitope core. A panel of point mutants expressing the ectodomain of A33 protein was generated and analyzed by either binding assays such as ELISA and immunoprecipitation or a functional assessment by blocking MAb-1G10 mediated comet inhibition in cell culture.
These results confirm L118 as a component of the MAb-1G10 binding epitope, and further identify D115 as an essential residue. By defining the minimum conformational structure, as well as the conformational arrangement of a short peptide sequence recognized by MAb-1G10, these results introduce the possibility of designing small molecule mimetics that may interfere with the function of A33 in vivo. This information will also be useful for designing improved assays to evaluate the potency of monoclonal and polyclonal products that target A33 or A33-modulated EV dissemination.
A33 是一种 II 型整合膜蛋白,表达于痘苗病毒(VACV)的细胞外包膜形式上。用 A33 定向单克隆抗体被动转移或用 A33 亚单位疫苗接种可在动物模型中提供针对致死性痘病毒攻击的保护。A33 的同源物在正痘病毒属的成员中高度保守,是在疫苗或针对细胞外包膜病毒活性的检测中包含的潜在候选物。一种针对 VACV A33 的单克隆抗体 MAb-1G10 已被证明针对构象依赖性表位。有趣的是,虽然它识别 VACV A33 以及相应的天花病毒同源物,但它不与猴痘病毒同源物结合。在这项研究中,我们利用随机噬菌体展示文库来研究 MAb-1G10 识别的对于促进痘苗病毒细胞间传播至关重要的表位。
通过用线性或构象随机噬菌体文库筛选,我们发现与 MAb-1G10 结合的噬菌体显示出共识基序 CEPLC,其中两个半胱氨酸残基之间形成二硫键,这对于 MAb-1G10 结合是必需的。尽管噬菌体基序不包含与 VACV A33 同源的线性序列,但结构建模和分析表明残基 D115 对于形成最小表位核心很重要。生成了表达 A33 蛋白外域的一组点突变体,并通过 ELISA 和免疫沉淀等结合测定或通过阻断 MAb-1G10 介导的细胞培养中彗星抑制作用来进行功能评估来进行分析。
这些结果证实 L118 是 MAb-1G10 结合表位的组成部分,并进一步确定 D115 是必需残基。通过定义最小构象结构以及 MAb-1G10 识别的短肽序列的构象排列,这些结果提出了设计可能干扰 A33 在体内功能的小分子模拟物的可能性。这些信息对于设计改进的测定法也将是有用的,该测定法可评估针对 A33 或 A33 调制的 EV 传播的单克隆和多克隆产品的效力。