Department of Earth and Environmental Science, University of Pennsylvania, Philadelphia, Pennsylvania, USA.
PLoS One. 2012;7(10):e45712. doi: 10.1371/journal.pone.0045712. Epub 2012 Oct 15.
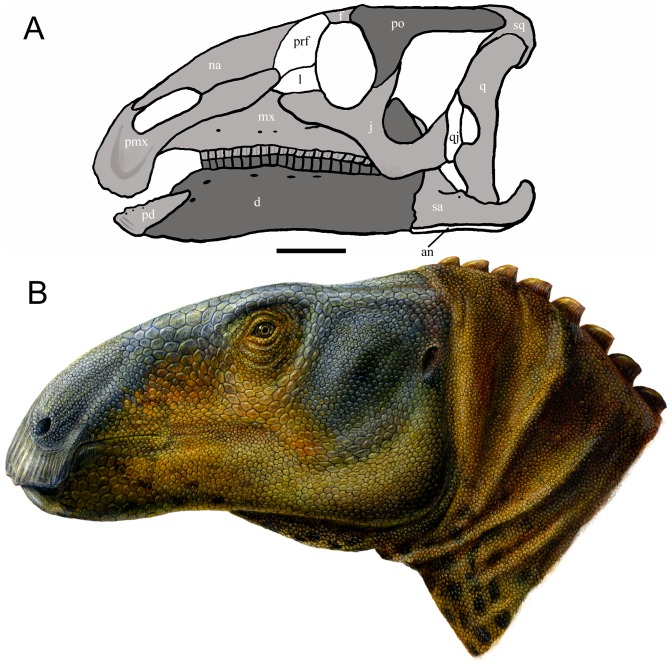
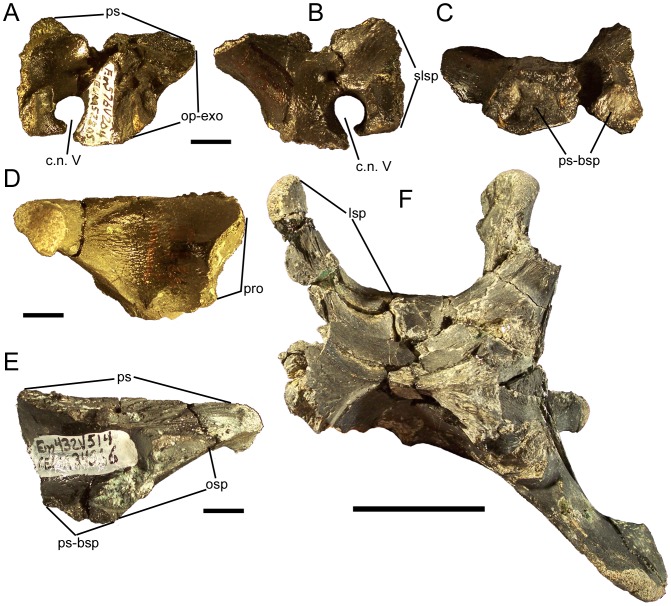
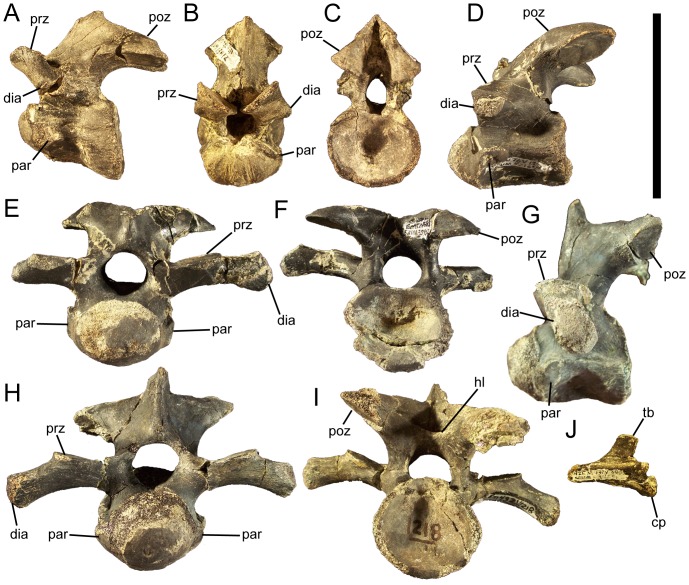
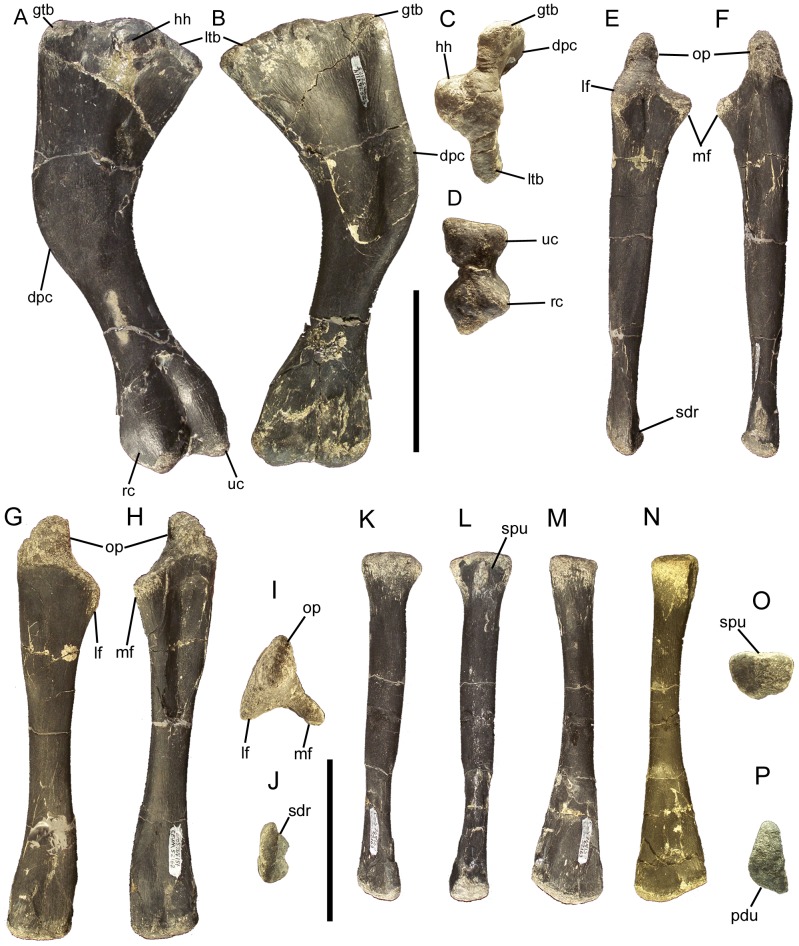
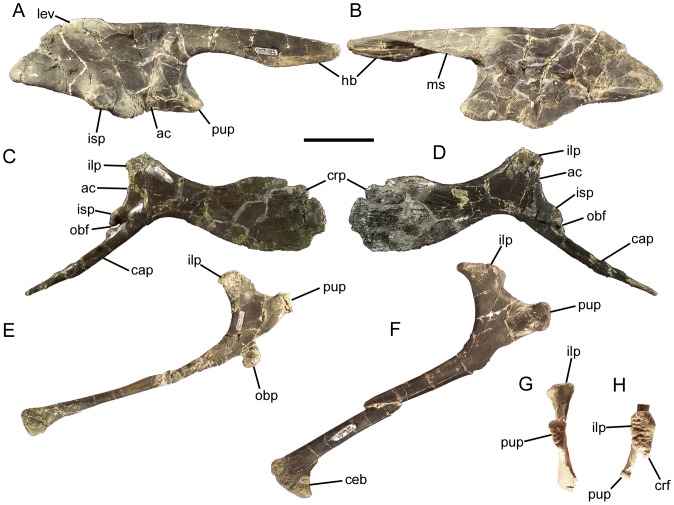
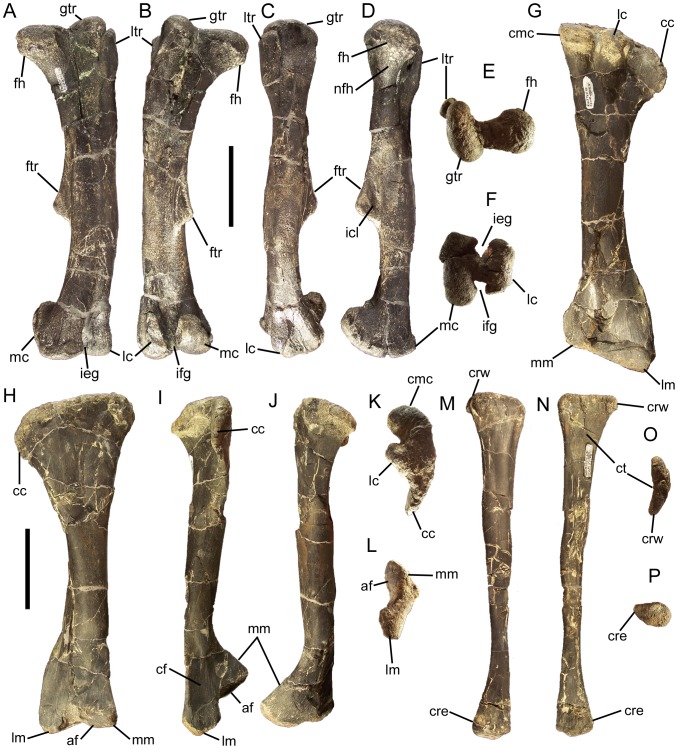
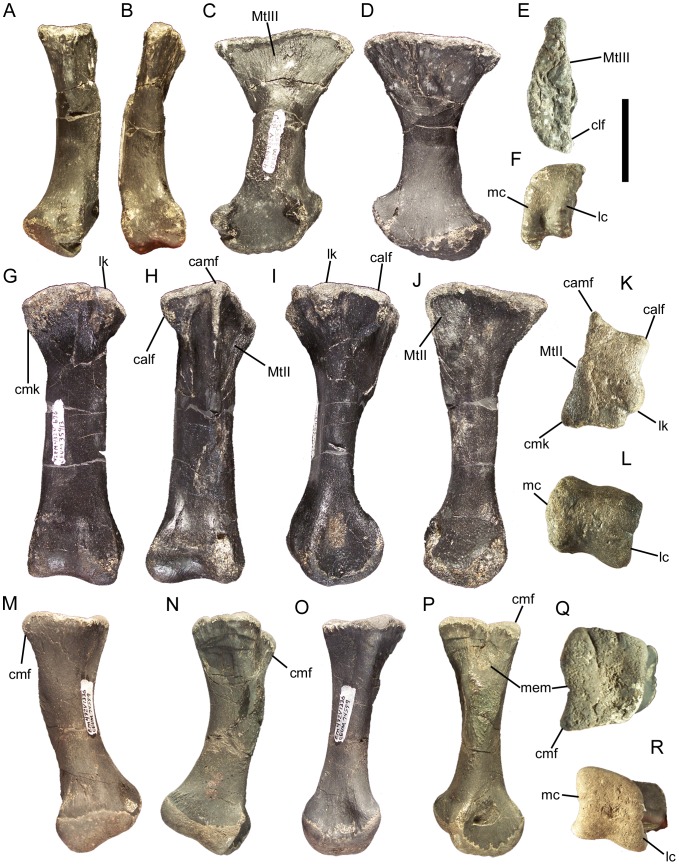
Eolambia caroljonesa is known from copious remains from the lower Cenomanian Mussentuchit Member of the Cedar Mountain Formation in eastern Utah; however, the taxon has been only briefly described. Thus, we present herein a complete osteological description of Eolambia.
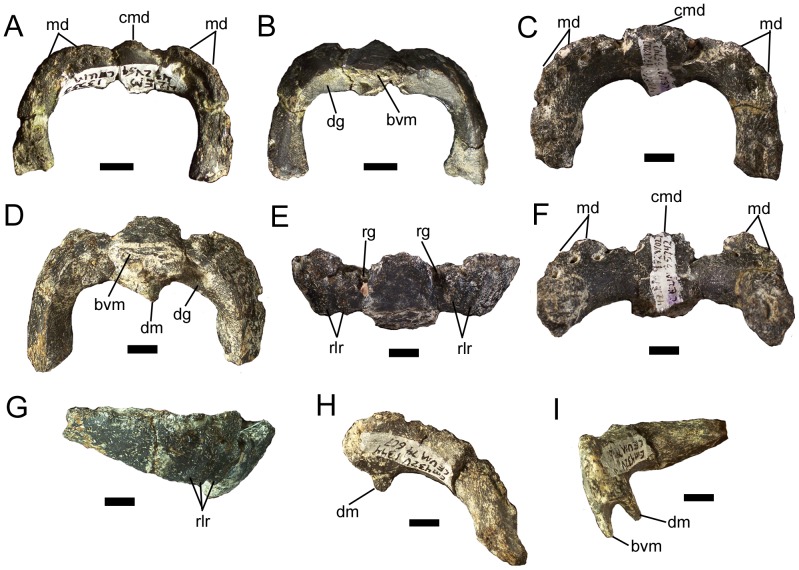
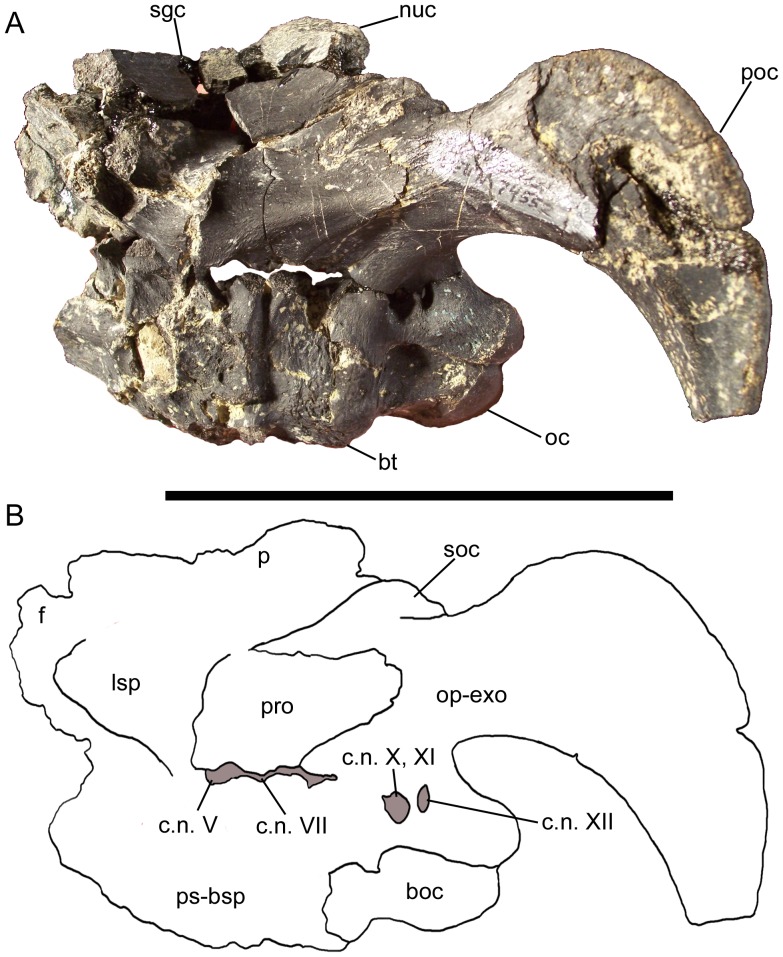
METHODOLOGY/PRINCIPAL FINDINGS: The description of Eolambia presented here is based upon the holotype partial skeleton (CEUM 9758), paratype partial skull (CEUM 5212), and abundant disarticulated elements from two bonebeds that contain juvenile individuals. These remains allow the skeletal anatomy of Eolambia to be documented almost fully and a revised diagnosis to be proposed.
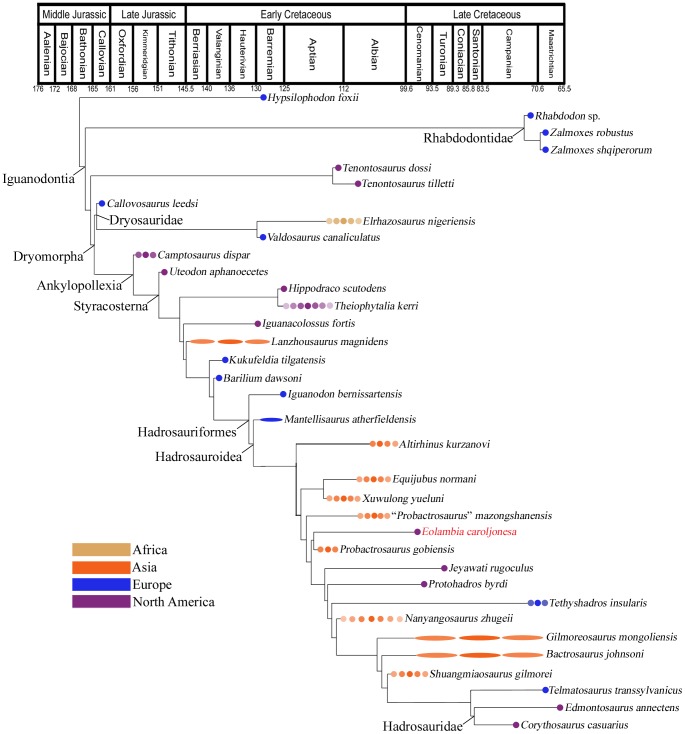
CONCLUSIONS/SIGNIFICANCE: The description provided here facilitates comparisons between Eolambia and other iguanodontians and allows Eolambia to be coded for additional characters in phylogenetic analyses. The close affinity between Eolambia and Probactrosaurus gobiensis from the Early Cretaceous of China supports previous hypotheses of faunal interchange between Asia and North America in the early Late Cretaceous.
伊奥兰巴·卡罗利纳莎(Eolambia caroljonesa)的化石记录丰富,来自于犹他州东部下白垩统雪松山组的穆斯廷图奇特阶;然而,该种仅被简要描述过。因此,我们在此提供了伊奥兰巴的完整骨骼描述。
方法/主要发现:本文对伊奥兰巴的描述基于模式标本(CEUM 9758)的部分骨骼、副型标本(CEUM 5212)的部分头骨,以及两个含幼年个体的骨床中大量分离的骨骼元素。这些遗骸使伊奥兰巴的骨骼解剖结构得以几乎完整地记录下来,并提出了一个修订后的诊断。
结论/意义:这里提供的描述有助于在伊奥兰巴和其他禽龙类之间进行比较,并允许在系统发育分析中为伊奥兰巴编码更多的特征。伊奥兰巴与来自中国早白垩世的戈壁原巴克龙(Probactrosaurus gobiensis)之间的密切亲缘关系,支持了早期晚白垩世亚洲和北美的动物群交流假说。