Division of Biological Sciences, Graduate School of Science, Nagoya University, Nagoya, Aichi 464-8602 Japan.
Mol Biol Cell. 2013 Jun;24(11):1765-75, S1-8. doi: 10.1091/mbc.E12-12-0907. Epub 2013 Mar 27.
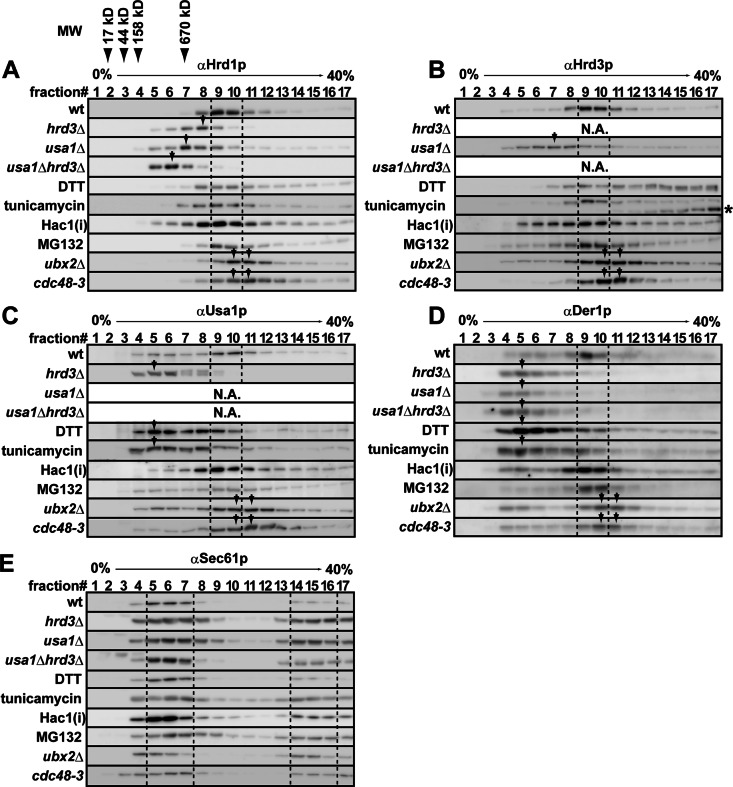
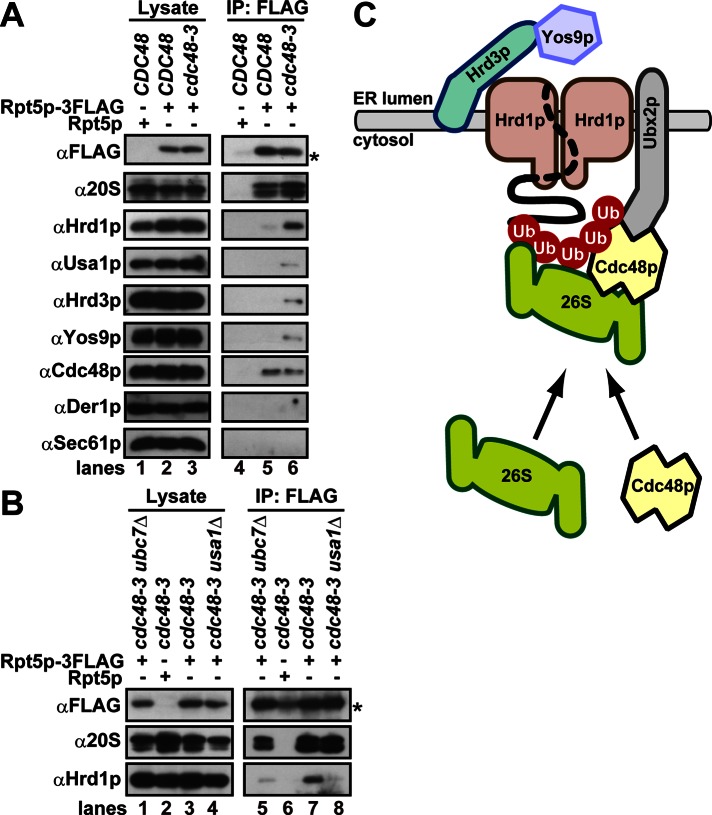
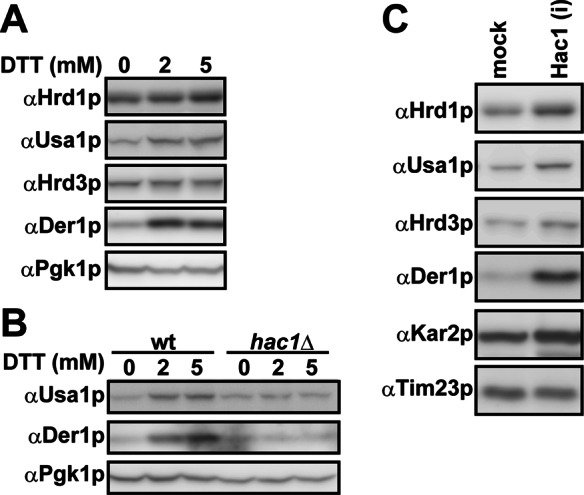
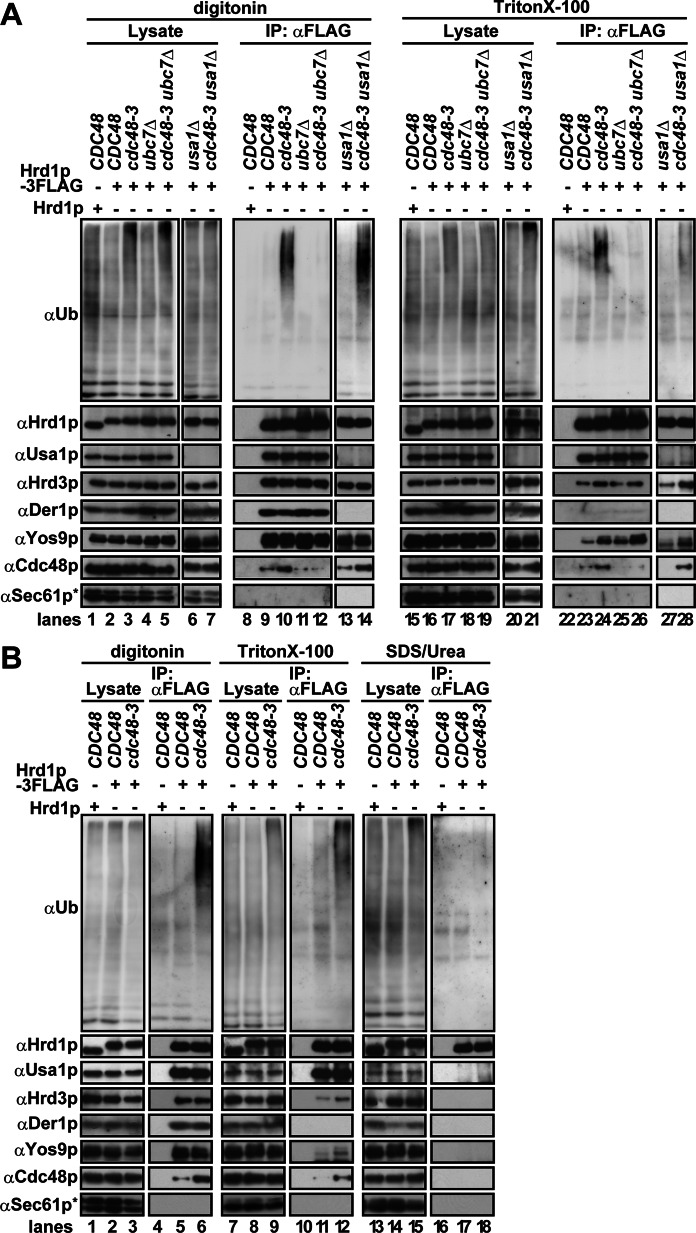
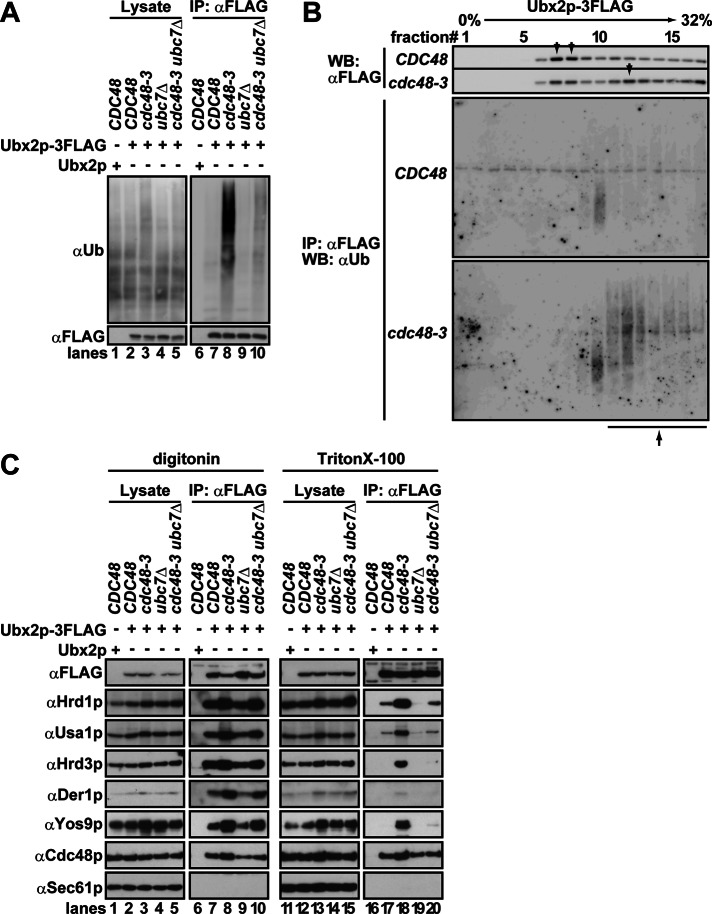
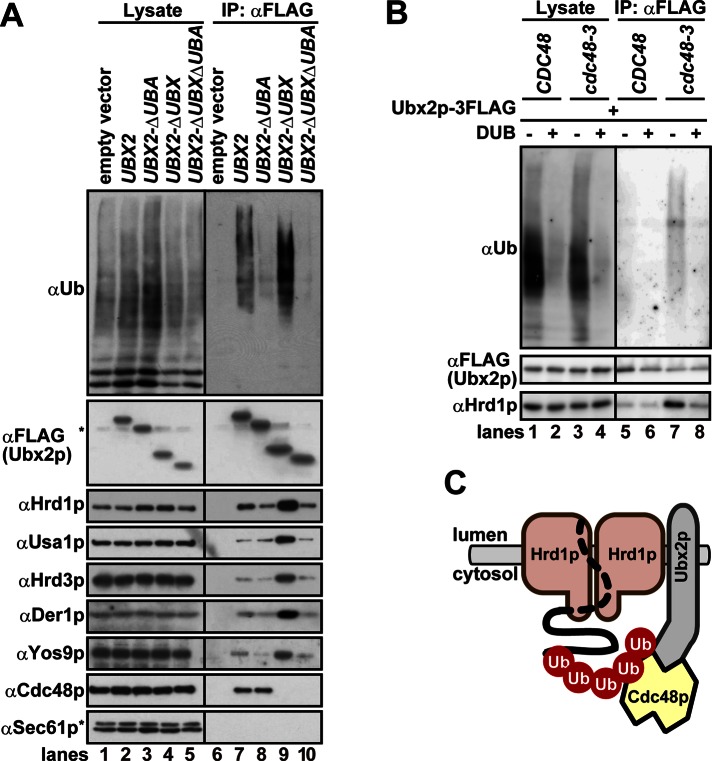
During endoplasmic reticulum-associated degradation (ERAD), misfolded lumenal and membrane proteins in the ER are recognized by the transmembrane Hrd1 ubiquitin ligase complex and retrotranslocated to the cytosol for ubiquitination and degradation. Although substrates are believed to be delivered to the proteasome only after the ATPase Cdc48p/p97 acts, there is limited knowledge about how the Hrd1 complex coordinates with Cdc48p/p97 and the proteasome to orchestrate substrate recognition and degradation. Here we provide evidence that inactivation of Cdc48p/p97 stalls retrotranslocation and triggers formation of a complex that contains the 26S proteasome, Cdc48p/p97, ubiquitinated substrates, select components of the Hrd1 complex, and the lumenal recognition factor, Yos9p. We propose that the actions of Cdc48p/p97 and the proteasome are tightly coupled during ERAD. Our data also support a model in which the Hrd1 complex links substrate recognition and degradation on opposite sides of the ER membrane.
在内质网相关降解(ERAD)过程中,内质网中错误折叠的腔和膜蛋白被跨膜 Hrd1 泛素连接酶复合物识别,并被反向转运到细胞质中进行泛素化和降解。尽管人们认为底物只有在 ATP 酶 Cdc48p/p97 作用后才能递送到蛋白酶体,但关于 Hrd1 复合物如何与 Cdc48p/p97 和蛋白酶体协调以协调底物识别和降解的知识有限。在这里,我们提供的证据表明,Cdc48p/p97 的失活会使反向转运停滞,并触发包含 26S 蛋白酶体、Cdc48p/p97、泛素化底物、Hrd1 复合物的选定成分和腔识别因子 Yos9p 的复合物的形成。我们提出,在 ERAD 过程中,Cdc48p/p97 和蛋白酶体的作用紧密耦合。我们的数据还支持这样一种模型,即 Hrd1 复合物在 ER 膜的相对侧连接底物识别和降解。